Sunishtha Singh Yadav1, Manish Kumar Singh2 and Rajesh Singh Yadav3*
1Amity Institute of Biotechnology, Amity University, Noida - 201 301 (UP), India
2Department of Pharmacology, King George Medical University, Lucknow 226 003, UP, India
3Department of Criminology and Forensic Science, School of Applied Sciences, Dr. Harisingh Gour Central University, Sagar - 470 003 (MP), India
*Corresponding Author:
Rajesh Singh Yadav
Department of Criminology and Forensic Science, School of Applied Sciences
Dr. Harisingh Gour Central University
Sagar – 470 003 (MP), India
Tel: +91-7582-264122
Fax: +91-7582-264163
E-mail: razitrc@gmail.com
Received date: December 28, 2015; Accepted date: January 13, 2016; Published date: January 22, 2016
Citation: Yadav SS, Singh MK, Yadav RS. Organophosphates Induced Alzheimer’s Disease: An Epigenetic Aspect. J Clin Epigenet. 2016, 2:1. DOI: 10.21767/2472-1158.100010
Copyright: © 2016 Yadav RS, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Keywords
Organophosphates; Cholinergic deficits; Alzheimer’s disease; Mitochondrial dysfunctions; Apoptosis
Introduction
The lack of full proof therapy for neurodegenerative disorders and CNS diseases resulted in an increase negative impact on the quality of life and become an economic burden onto the society. Alzheimer’s disease is a progressive neurodegenerative disorder associated with the loss of cholinergic neurons and the presence of excessive neuritic plaques containing amyloid β protein and abnormal tau protein filaments in the form of neurofibrillary tangles [1,2]. Decreased level of acetylcholine in Alzheimer’s disease patients appears to be a critical element in producing dementia and memory disorders [3]. Cholinergic neurodegeneration have also been found to be associated with the loss of acetylcholine [4]. The disease includes multiple pathophysiological factors, including flawed amyloid beta protein metabolism, abnormalities of glutamatergic, adrenergic, serotonergic and dopaminergic neurotransmission and also the involvement of inflammatory cytokines [1,5]. Although the Alzheimer’s disease is an age related progressive neurological disorder, early onset of disease in the 40s and 50s has also been reported in people [6]. Several factors such as environmental exposure and genetic predisposition could be involve [7,8] in reducing the synthesis of acetylcholine, deposition of amyloid beta [9] alterations in tau protein in the brain [10], oxidative stress and inflammatory cytokines [11]. The disease is rapidly progressive, leading to the loss of cognitive functions, especially memory processes and associated behavior. The precise cause of Alzheimer’s disease is not entirely known, but the role of genetic factors, including amyloid precursor protein (APP), presenilin 1, presenilin 2 and apolipoprotein E gene has been found to be associated with the pathophysiology of Alzheimer’s disease [12]. The prevalence of Alzheimer’s disease due to genetic susceptibility in the pesticide exposed population becomes a serious matter of concern in recent years [5,13].
The production and consumption of pesticides have increased many folds after the Green Revolution in India and many other parts of the globe. Most of these pesticides are neurotoxic in nature; their direct or indirect exposure to human may lead to the neurological deformities and disorders. Of various insecticides, organophosphates are frequently used in agriculture, homes and public health programme to control pests and vectors and in veterinary practices to control ectoparasites, hence distributed in the environment [14,15]. Human exposure to these organophosphates is quite imminent due to the indiscriminate and excessive use in occupational and non-occupational settings [16,17] which may associate with various neurodegenerative diseases [5,18,19]. Cases of organophosphate poisoning from India and many other countries have been frequently reported [15,20]. Also, these organophosphate compounds are expected to involve in the highest incidences of suicidal poisoning in India [15,21]. High levels of residues of organophosphate and their metabolites detected in the dietary products and biological tissues of exposed individuals are again a matter of concern due to associated toxic health effects [22-24]. The possible role of pesticide exposure, especially organophosphates in Alzheimer’s disease and dementia has not received large attention and therefore its consequences occur in the society [25]. The present review has therefore been focused on organophosphates induced mitochondrial dysfunctions, apoptotic signaling, amyloid processing, β-amyloid segregation and impairment of synaptic transmission to establish a possible epigenetic link between the pesticide exposure and pathophysiology of Alzheimer’s disease and clinical implications.
Organophosphates Induced Neurotoxicity
The possible association of chronic pesticide exposure with the increased prevalence of dementia and Alzheimer’s disease has been reported [13,26]. Organophosphates severely damage the brain and affect the behavioral pattern and neurological activities in exposed individuals [27-29]. The specific pattern of damage is cholinergic dysfunctions associated with learning and cognitive deficits, impaired neurobehavioral and neuropsychological performance [20]. Human exposure to organophosphate including monocrotophos, chlorpyrifos and dichlorvos is extensively reported due to their wide applications and presence as a contaminant in the dietary and food products [15,22]. Study on farmers in Egypt using organophosphates, including monocrotophos reported that 50% of the workers had neurological symptoms such as loss of reflexes [30]. The workers, including applicators, technicians, and engineers working in Egyptian cotton production have been found to have a substantially higher degree of chlorpyrifos exposure associated with neurobehavioral deficits [31]. They further demonstrated a dose effect relationship between urinary trichloro-2-pyridinol (TCPy), a biomarker for chlorpyrifos exposure and both plasma butyl cholinesterase (BChE) and red blood cell AChE in chlorpyrifos exposed workers [17]. In another study, [32] reported that chlorpyrifos can result in persistent alterations in axonal transport in the living mammalian brain which may lead to neurological deficits in humans repeatedly exposed to organophosphates. [33], in his study suggested that exposure through different organophosphorus pesticides could lead to the cognitive, psychomotor and emotional disturbances in individuals. The presence of residues of organophosphates, including chlorpyrifos and its metabolites in maternal prenatal and postnatal blood, cord blood and in maternal and child urine has suggested the developmental neurotoxicity of organophosphates [34-36]. In a recent study, [37] reported that exposure to phosphomedon in rats caused neurobehavioral abnormalities such as reduce food intake, weight loss, increase water intake, low defecation frequency and altered locomotion frequency. At the same time a severe histopathological changes were also observed, which was found to be associated with the neurobehavioral changes suggesting the neuronal loss. In the environment, the risk of exposure through multiple chemicals at a single time may cause their synergistic effects. In this connection a study on the exposure of real life doses of malathion, DEET and permethrin, alone or in combination in rats have been reported to cause significant neurobehavioral deficits and neuronal degeneration [38]. Repeated dose of malathion and diazinon in rats has been found to cause oxidative stress, inhibit brain and plasma cholinesterase cause, histopathological and immune alterations in brain and other body organs [39]. Recently, [40] showed that combined exposure of chlorpyrifos and lead acetate reduce the activities of brain antioxidant enzymes and AChE and increased lipid peroxidation. The changes were further linked to the altered histopathological structure of the cerebral cortex in rats [41] found that diazinon and its oxygen metabolite diazoxon increases oxidative stress in astrocytes and adversely affect astrocyte function, resulting in inhibited neurite outgrowth in hippocampal neurons linked to the decreased levels of astrocytic fibronectin.
Organophosphate Induced Oxidative Stress and Alzheimer's Disease
Organophosphates induced free radical generation linked with enhanced oxidative stress in humans has been suggested as one of the key mechanism of their neurotoxic alterations [42,43]. The amyloid β protein is found to be an important factor to enhance oxidative stress linked with increased levels of lipid peroxidation products including malondialdehyde, 4-hydroxynonenal (HNE) and acrolein [44,45]. These toxic products, formed as a result of oxidative stress alter the cellular structure and physiological function of the brain and leads to neurodegenerative diseases, including Alzheimer's disease [46,47]. The involvement of lipids, inflammatory mediators in the production and accumulation of β-amyloid and enhanced oxidative stress in Alzheimer’s disease has also been reported [48-50]. We have also reviewed and suggested that the generation of reactive oxygen and nitrogen species as a result of pesticide exposure could damage the lipid membrane and alter the composition of lipid rafts leading to various brain related disorders [51]. On the other hand the accumulation of transition metals including iron further involve in the generation of free radicals through the process of Fenton’s reaction. Docosahexenoic (DHA), dietary essential polyunsaturated fatty acids (PUFA), another target in oxidative damage has been found to link with the cognitive decline and neuronal dysfunction in Alzheimer's disease [52]. The alterations in the brain lipid profile, including phospholipids, sphingomylein, ceramide and ganglioside could modulate the signaling cascade and neural function, leading to neurological disorders, including Alzheimer’s disease [53,54]. Further altered levels of sphingomyelins andceramides in Alzheimer’s disease brains have been reported as a result of sphingomyelin hydrolysis [55].
Organophosphate Induced Neuronal Loss via Apoptosis
Acetyl cholinesterase (AChE), an enzyme involved in the synaptic transmission is the prime target of action of organophosphates. They inhibit the activity of AChE in an irreversible manner and caused over accumulation of the levels of acetylcholine at the synaptic junction leading to desensitization of receptors and finally paralysis and cell death [49,56]. Mitochondrial dysfunction and oxidative metabolism are considered to be the key mechanism for organophosphates induced apoptosis and in the pathogenesis of Alzheimer's disease [57]. Mitochondria play a vital role in apoptotic pathways, as it contains decisive apoptotic factors, including cytochrome C in their intermembranous space [58]. Once the cytochrome C release into the cytosol, it initiates the activation of caspase-cascade mechanisms of apoptosis [59]. The anti-apoptotic protein family, such as Bcl- 2 and Bcl-xL strictly regulate the release of cytochrome C and maintain the ratio between Bcl2/Bax [60,61]. The decrease in the ratio of Bcl2/Bax due to oxidative stress initiates the release of cytochrome C and activation of caspase-cascade and leads to the apoptosis [62]. The activation of caspases provides a crucial factor in the implementation of mitochondria mediated apoptosis [63]. Enhanced oxidative stress following exposure to monocrotophos in rats has been found to affect mitochondrial complex I, II and IV associated with decreased production of ATP [64]. The over activation of apoptotic factors in central nervous system can contribute to the neuronal cell death and may cause neurodegenerative disorders such as Alzheimer’s and Parkinson’s diseases [65,66]. Several studies have suggested that the toxicological response of organophosphates and its compounds may cause neuronal apoptosis linked with organophosphate induced delayed neuropathy (OPIDN) [58,67].
The alteration in lipid rafts composition may also initiate the neurodegeneration and apoptosis through various pathways including aggregation of amyloid beta. Increased expression of apoptotic proteins including Bax, JNK, c-jun, ERK1/2, MAP kinases and decreased expression of anti-apoptotic proteins such as p38 MAP kinase, Bcl-2, Bcl-xL have been reported in the organophosphate induced neurotoxicity [60,61,68] reported that exposure to chlorpyrifos may induce apoptosis in primary cortical neurons cultured from embryonic day 17 or newborn rats independently of AChE inhibition. They further suggested that the activation of the ERK1/2 and JNK MAP kinases involve in apoptotic and activation of the p38 MAP kinase in antiapoptotic mechanism in cortical neurons following exposure to chlorpyrifos. Exposure to organophosphates enhanced the levels of intracellular calcium, which triggered the activation of calpains in nerve tissues [69]. This activated calpains may further activate the cyclin-dependent kinase 5 (Cdk5) and involved in the neuronal cell death [59,70]. The detailed mechanism of neuronal apoptosis is illustrated in the Figure 1. Exposure to organophosphates, including monocrotophos, dichlorvos, chlorfenvinphos, chlorpyrifos, malathion, quinalphos etc. have been found to disrupt the balance of antioxidant and pro-oxidant in the brain and linked with enhanced oxidative stress [51,71]. Increased lipid peroxidation in brain regions and cerebro-spinal fluid of rats has been reported following exposure to malathion [72]. Exposure of triazophos in rats has been found to cause increased lipid peroxidation associated with decreased mRNA and protein expression of brain derived neurotrophic factor (BDNF) and reduced glutathione in hippocampus suggesting the role of oxidative stress in the toxicity of triazophos [73]. Further evidences also indicated that the generation of reactive nitrogen species through activated astrocytes and oxidative stress are involved in various neurodegenerative diseases, including Alzheimer’s disease, ischemia, epilepsy etc. [74-76].
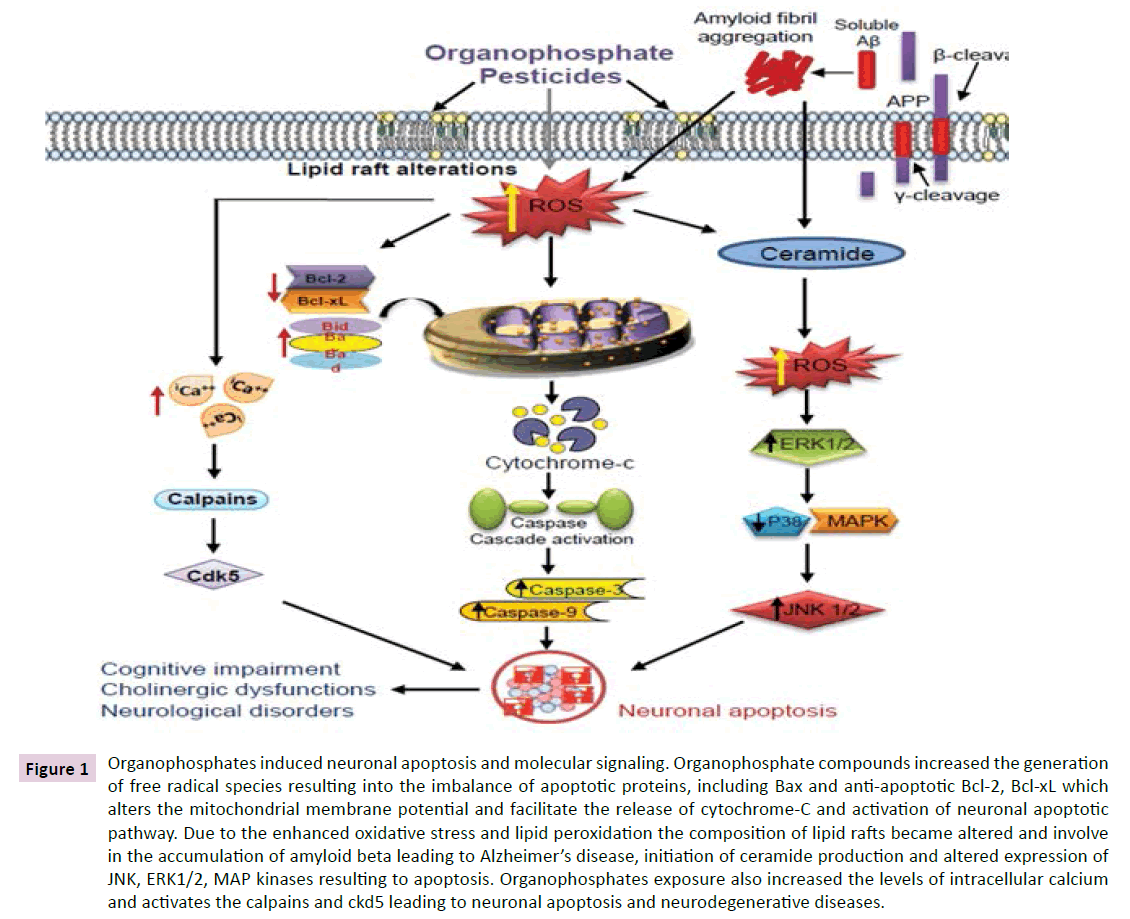
Figure 1 Organophosphates induced neuronal apoptosis and molecular signaling. Organophosphate compounds increased the generation of free radical species resulting into the imbalance of apoptotic proteins, including Bax and anti-apoptotic Bcl-2, Bcl-xL which alters the mitochondrial membrane potential and facilitate the release of cytochrome-C and activation of neuronal apoptotic pathway. Due to the enhanced oxidative stress and lipid peroxidation the composition of lipid rafts became altered and involve in the accumulation of amyloid beta leading to Alzheimer’s disease, initiation of ceramide production and altered expression of JNK, ERK1/2, MAP kinases resulting to apoptosis. Organophosphates exposure also increased the levels of intracellular calcium and activates the calpains and ckd5 leading to neuronal apoptosis and neurodegenerative diseases.
Organophosphate induced Inflammation and Alzheimer’s disease
The role of inflammation in the etiology and pathogenesis of Alzheimer’s disease has been suggested [77,78]. The activation of these inflammatory cytokines occurs due to the enhanced oxidative stress, which may involve in the process of neurodegeneration in Alzheimer’s diseases [79]. The role of microglia in Alzheimer’s disease has been suggested due to the presence of plaque associated microglia that exhibits a reactive phenotype [77,80]. The inflammatory response is primarily expressed by the activation of glial cells, macrophages and oligodendrocytes in the brain associated with the triggering of pro-inflammatory cytokines including interleukin (IL)-1β, IL-18 and IL-33 and linked with the infection, autoimmunity, neuroinflammation and associated disorders [81-83]. These activated microglias are involved in the process of apoptosis and neuronal death via the secretion of various proinflammatory molecules and cytokines (IL-1, IL-6 and TNF-α) and also facilitate the production and deposition of amyloid in the brain [77,84,85] have demonstrated that dichlorvos exposure in rats can activate microglial cells and cause apoptosis through the upregulation of pro-inflammatory molecules like nitric oxide, TNF-α, and IL-1β. The microglial apoptosis has also been found to be associated with the increased expression of Bax in mitochondria, cytochrome c release from mitochondria, and caspase-3 activation. The role of inflammasomes in the pathophysiology of neuroinflammation and neurodegenerative diseases including dementia, memory and cognitive dysfunctions has been reported in the last decades [78,83,86]. Also the role of inflammasomes in the etiologies of Alzheimer’s disease has been suggested [87]. The roles of DNA methylation and hydroxymethylation in the development and potential treatment of AD have also been reported [88,89] in their review has suggested that oxidative stress, neuroinflammation, microtubule alterations, synthesis of beta amyloid, calcium dyshomeostasis and mitochondrial dysfunction all are contributing factors in organophosphate induced neurological diseases.
Prevention and Suggestions
The risk of human exposure to organophosphates is enhanced several times in developing countries due to the irregularities in safety measures. Due to the high risk of neurotoxic impact of these organophosphate compounds on human health, especially on developing children, it deserves the attention of regulatory agencies and prevention authorities. There is a need to develop the protective measures for agricultural workers and other individuals occupationally exposed to these pesticides. The industrial manufacturers should have to use proper safety measures and also aware the workers, users and general public about their harmful consequences through improper handling and uses. There should be trainings and workshops on these pesticides, including organophosphates to awareness and educate the users in agricultural sectors and public health programs. The research should be continued in the area of developing substitute of these toxic compounds for their use in agriculture and public health. Specific biomarkers could be identified in the study of molecular mechanisms of neurotoxicity of organophosphate compounds through parallel studies in humans and animals which could help to develop a protective and effective cure [90]. At the same time these biomarkers may also provide strategies to identify the risk in exposed individuals. In recent years natural and pharmacological agents have been found to combat the neurotoxic effects of organophosphate compounds [91-95]. Further, to assess the neurotoxic impact of specific pesticides in human populations, there is a need to perform well designed epidemiological studies which could provide useful information to research scientist working in the area of occupational, environmental and human health. Also, animal and alternate animal model research must be going on to find out the mechanisms of neurotoxicity of these pesticides at low dose levels comparable with real world exposure. Invasive research on molecular basis may improve the understanding of mechanism of neurotoxicity from organophosphate exposure and hence will be useful to develop protective measures.
Conclusion
The use of pesticides, especially organophosphates and its associated neurochemical alterations and neurological disorders in both adults and children become a serious concern among the health scientists to use protective and preventive approaches for minimizing its neurotoxicity. Some of the organophosphates are banned in India and in several other countries, but their injudicious use is further a matter of concern world over. The present review may help to understand the detailed mechanism of organophosphate induced neurotoxicity and to find out preventive measures accordingly. The study may provide new insights into neurotoxicity and open new vistas for regulatory agencies to use the human data in risk assessments of pesticides. In the present study, an attempt has been made to add new information to the literature regarding the overall neurotoxicity and the general mechanisms of toxic actions of organophosphate pesticides. A detailed understanding of the molecular and cellular toxic events linked with signaling cascade in the brain is required for effective cure, prevention and management of organophosphate induced neurotoxicity.
Conflict of Interest
There is no conflict of interest.
Acknowledgement
The authors are thankful to Dr. Harisingh Gour Central University, Sagar (MP), India for providing the opportunity to work and their support and interest. Dr. Rajesh Singh Yadav is also thankful to the University Grants Commission (UGC), New Delhi, India for providing the UGC-BSR startup research grants.
References
- Breunig JJ, Guillot-Sestier MV, Town T (2013) Brain injury, neuroinflammation and Alzheimer's disease. Front Aging Neurosci 5:26.
- Lord J, Cruchaga C (2014) The epigenetic landscape of Alzheimer's disease. Nature Neuroscience 17: 1138–1140.
- Nagele RG, D'Andrea MR, Anderson WJ, Wang HY (2002) Intracellular accumulation of beta-amyloid(1-42) in neurons is facilitated by the alpha 7 nicotinic acetylcholine receptor in Alzheimer's disease. Neuroscience 110:199-211.
- Hamlin AS, Windels F, Boskovic Z, Sah P, Coulson EJ (2013) Lesions of the basal forebrain cholinergic system in mice disrupt idiothetic navigation. PLoS One 8:e53472.
- Wingo TS, Rosen A, Cutler DJ, Lah JJ, Levey AI (2012) Paraoxonase-1 polymorphisms in Alzheimer's disease, Parkinson's disease, and AD-PD spectrum diseases. Neurobiol Aging 33:e13-e5.
- Kim J, Ahn H, Han BC, Lee SH, Cho YW, Kim CH et al (2014) Korean red ginseng extracts inhibit NLRP3 and AIM2 inflammasome activation. ImmunolLett 158: 143–150.
- Bertram L, Tanzi RE (2009) Genome wide association studies in Alzheimer’s disease. Hum. Mol. Genet 18: R137-R45.
- Kamboh M, Demirci F, Wang X, Minster R, Carrasquillo M, Pankratz V et al (2012) Genome wide association study of Alzheimer’s disease. Transl. Psychiatry 2:e117.
- Palop JJ, Mucke L (2010) Amyloid β induced neuronal dysfunction in Alzheimer’s disease: from synapses toward neural networks. Nat Neurosci 13: 812–818.
- Ballatore C, Lee VMY, Trojanowski JQ (2007) Tau mediated neurodegeneration in Alzheimer’s disease and related disorders. Nat Rev Neurosci 8: 663–672.
- Agostinho PA, Cunha R, Oliveira C (2010) Neuroinflammation, oxidative stress and the pathogenesis of Alzheimer’s disease. Curr Pharm Des 16: 2766-2778
- Foraker J, Millard SP, Leong L, Thomson Z, Chen S, Keene CD, et al. (2015) The APOE Gene is Differentially Methylated in Alzheimer's Disease. J Alzheimers Dis 48:745-755.
- Campdelacreu J (2014) Parkinson’s disease and Alzheimer disease: environmental risk factors. Neurologia 29: 541-549.
- Eaton DL, Daroff RB, Autrup H, Bridges J, Buffler P, et al (2008) Review of the toxicology of chlorpyrifos with an emphasis on human exposure and neurodevelopment. Crit Rev Toxicol 38: 1–125.
- World Health Organization (2009) Health implications from monocrotophos use: a review of the evidence in India.
- Curl CL, Fenske RA, Kissel JC, Shirai JH, et al (2002) Evaluation of take-home organophosphorus pesticide exposure among agricultural workers and their children. Environ Health Perspect 110: A787–A792.
- Farahat FM, Ellison CA, Bonner MR, McGarrigle BP, Crane AL, Fenske RA, et al (2011) Biomarkers of chlorpyrifos exposure and effect in Egyptian cotton field workers. Environ Health Perspect 119:801-806.
- Wang A, Costello S, Cockburn M, Zhang X, Bronstein J, Ritz B (2011) Parkinson's disease risk from ambient exposure to pesticides. Eur J Epidemiol 26:547-555.
- Das K, Ghosh M, Nag C, Nandy SP, Banerjee M, Datta M, et al (2011) Role of Familial, Environmental and Occupational Factors in the Development of Parkinson's Disease. Neurodegener Dis 8:345-351.
- Roldan-Tapia L, Nieto-Escamez FA, Agulia EM, et al (2006) Neuropsychological sequelae from acute poisoning and long-term exposure to carbamate and organophosphate pesticides. Neurotoxicology and Teratology 28: 694-703.
- Tsai CY, Chang AY, Chan JY, Chan SH (2014) Activation of PI3K/Akt signaling in rostral ventrolateral medulla impairs brain stem cardiovascular regulation that underpins circulatory depression during mevinphos intoxication. BiochemPharmacol 88:75-85.
- Mathur HB, Agarwal HC, Sapna J, Nirmali S (2005) Analysis of Pesticide residues In Blood Samples From Villages of Punjab CSE/PML/PR-21/2005.
- Moreno-Banda G, Blanco-Munoz J, Lacasana M, Rothenberg SJ et al (2009) Maternal exposure to floricultural work during pregnancy, PON1 Q192R polymorphisms and the risk of low birth weight. Sci Total Environ 407:5478-5485.
- Srivastava S, Narvi SS, Prasad SC (2011) Levels of select organophosphates in human colostrum and mature milk samples in rural region of Faizabad district, Uttar Pradesh, India. Hum ExpToxicol 30:1458-1463.
- Sanborn M, Kerr KJ, Sanin LH, Cole DC, Bassil KL, Vakil C (2007) Non-cancer health effects of pesticides. Can. Fam. Physician 53: 1712–1720.
- Zaganas I, Kapetanaki S, Mastorodemos V, Kanavouras K et al (2013) Linking pesticide exposure and dementia: what is the evidence? Toxicology 10: 307-311.
- Rohlman DS, Anger WK, Lein PJ (2011) Correlating neurobehavioral performance with biomarkers of organophosphorous pesticide exposure. Neurotoxicology 32: 268–276.
- Burns CL, McIntosh LJ, Mink PJ, Jurek AM, Li AA (2013) Pesticide Exposure and Neurodevelopmental Outcomes: Review of the Epidemiologic and Animal Studies, Journal of Toxicology and Environmental Health, Part B: Critical Reviews 16:127-283.
- Berent S, Giordani B, Albers JW, Garabrant DH, Cohen SS, et al. (2014) Effects of occupational exposure to chlorpyrifos on neuropsychological function: A prospective longitudinal study. NeuroToxicology 41: 44–53.
- Amr M (1995) Neurological effects of pesticides. Study by the industrial medicine and occupational diseases, Faculty of Medicine, Cairo University (Reported in Pesticide News, December 30).
- Farahat FM, Fenske RA, Olson JR, Galvin K, Bonner MR, Rohlman DS, et al (2010) Chlorpyrifos exposures in Egyptian cotton field workers. Neurotoxicology 31:297-304.
- Hernandez CM, Beck WD, Naughton SX, Poddar I, Adam BL, et al (2015) Repeated exposure to chlorpyrifos leads to prolonged impairments of axonal transport in the living rodent brain. NeuroToxicology 47: 17–26
- Ismail AA, Bodner TE, Rohlman DS (2012) Neurobehavioral performance among agricultural workers and pesticide applicators: a meta-analytic study. Occup Environ Med 69:457–464.
- Eskenazi B, Marks, AR, Bradman A, Harley K, Barr DB, Johnson C, Morga N, Jewell NP (2007) Organophosphate pesticide exposure and neurodevelopment in young Mexican- American children. Environ Health Perspect 115:792-798.
- Barr DB, Ananth CV, Yan X, Lashley S, Smulian JC, et al. (2010) Pesticide concentrations in maternal and umbilical cord sera and their relation to birth outcomes in a population of pregnant women and newborns in New Jersey. Sci Total Environ 408: 790–795.
- Engel SM, Wetmur J, Chen J, Zhu, C, Barr DB, Canfield RL, Wolff MS (2011) Prenatal exposure to organophosphates, paraoxonase 1, and cognitive development in childhood. Environ Health Perspect 119: 1182–1188.
- Hazarika R (2014) Neurotoxic impact of organophosphate pesticide phosphomedon on the albino rat. J Environ Biol 35:427-30.
- Abdel-Rahman A, Dechkovskaia AM, Goldstein LB, Bullman SH, Khan W, El-Masry EM, Abou-Donia MB (2004) Neurological deficits induced by malathion, DEET, and permethrin, alone or in combination in adult rats. J Toxicol Environ Health A 67:331-356.
- Baconi DL, Barca M, Manda G, Ciobanu AM, Balalau C (2013) Investigation of the toxicity of some organophosphorus pesticides in a repeated dose study in rats. Rom J MorpholEmbryol 54:349-356.
- Akande MG, Aliu YO, Ambali SF, Ayo JO (2014) Taurine mitigates cognitive impairment induced by chronic co-exposure of male Wistar rats to chlorpyrifos and lead acetate. Environ ToxicolPharmacol 37:315-325.
- Pizzurro DM, Dao K, Costa LG (2014) Diazinon and diazoxon impair the ability of astrocytes to foster neurite outgrowth in primary hippocampal neurons. ToxicolApplPharmacol 274:372-382.
- Slotkin TA, Oliver CA, Seidler FJ (2005) Critical periods for the role of oxidative stress in the developmental neurotoxicity of chlorpyrifos and terbutaline, alone or in combination. Dev Brain Res 157:172–180.
- Soltaninejad K, Abdollahi M (2009) Current opinion on the science of organophosphate pesticides and toxic stress: a systematic review. Med SciMonit 15:RA75-RA90.
- Poli G, Schaur RJ (2000) 4-Hydroxynonenal in the pathomechanisms of oxidative stress. IUBMB Life 50: 315-321.
- Lovell MA, Xie C, et al (2000) Acrolein, a product of lipid peroxidation, inhibits glucose and glutamate uptake in primary neuronal cultures. Free RadicBiol Med 29: 714-720.
- Smith DG, Cappai R, Barnham KJ (2007) The redox chemistry of the Alzheimer's disease amyloid β peptide. BiochimicaetBiophysicaActa 1768:1976–1990.
- Hardas SS, Sultana R, Clark AM et al (2013) Oxidative modification of lipoic acid by HNE in Alzheimer’s disease brain. Redox Biology 1:80–85.
- Ariga T, McDonald MP, Yu RK (2008) Role of ganglioside metabolism in the pathogenesis of Alzheimer’s disease-a review. Journal of Lipid Research 49:1157-1175.
- Sankhwar ML, Yadav RS, Shukla RK et al (2012) Impaired cholinergic mechanisms following exposure to monocrotophos in young rats. Hum ExpToxicol 31:606–616
- Zuo L, Hemmelgarn BT, Chuang CC, Best TM (2015) The Role of Oxidative Stress-Induced Epigenetic Alterations in Amyloid- β Production in Alzheimer's Disease. Oxid Med Cell Longev 604658.
- Sankhwar ML, Yadav RS, Shukla RK, Singh D, Ansari RW, et al (2013) Monocrotophos induced oxidative stress and alterations in brain dopamine and serotonin receptors in young rats. ToxicolInd Health.
- Lukiw WJ, Bazan NG (2008) Docosahexaenoic Acid and the Aging Brain. J. Nutr 138: 2510–2514.
- Jana A, Hogan EL, Pahan K (2009) Ceramide and neurodegeneration: Susceptibility of neurons and oligodendrocytes to cell damage and death. J NeurolSci278: 5–15.
- Haughey NJ, Bandaru VVR, Bai M, Mattson MM (2010) Roles for Dysfunctional Sphingolipid Metabolism in Alzheimer’s Disease Neuropathogenesis. BiochimBiophysActa 1801:878–886.
- He X, Huang Y, Li B, Gong CX, Schuchman EH (2010) Deregulation of sphingolipid metabolism in Alzheimer’s disease. Neurobiol Aging 31:398-408.
- Kobayashi H, Suzuki T, Sakamoto M, Hashimoto W et al (2007) Brain regional acetylcholinesterase activity and muscarinic acetylcholine receptors in rats after repeated administration of cholinesterase inhibitors and its withdrawl. Toxicology and Applied Pharmacology 219: 151–161.
- Kroemer G, Galluzzi L, Brenner C (2007) Mitochondrial membrane permeabilization in cell death. Physiol Rev 87:99–163.
- Li Y, Lein PJ, Liu C, Bruun DA, Tewolde T, Ford G, Ford BD (2011) Spatiotemporal pattern of neuronal injury induced by DFP in rats: a model for delayed neuronal cell death following acute OP intoxication. Toxicol. Appl. Pharmacol 253: 261–269.
- Wang YP, Mou DL, Song JF, Rao ZR, Li D, Ju G (2006) Aberrant activation of CDK5 is involved in the pathogenesis of OPIDN. J Neurochem 99:186–197.
- Cory S, Adams JM (2002) The Bcl2 family: regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2: 647-656.
- Edlich F, Banerjee S, Suzuki M, Cleland MM, Arnoult D, et al (2011) Bclx( L)retrotranslocatesBax from the mitochondria into the cytosol. Cell 145:104–116.
- Kashyap MP, Singh AK, Siddiqui MA, Kumar V, Tripathi VK, et al (2010) Caspase cascade regulated mitochondria mediated apoptosis in monocrotophos exposed PC12 cells. Chem Res Toxicol 23: 1663–1672.
- Paris C, Bertoglio J, Breard J (2007) Lysosomal and mitochondrial pathways in miltefosine induced apoptosis in U937 cells. Apoptosis 12:1257–1267.
- Masoud A, Kiran R, Sandhir R (2009) Impaired mitochondrial functions in organophosphate induced delayed neuropathy in rats. Cell MolNeurobiol 29:1245-1255.
- Hickey M, Chesselet MF (2003) Apoptosis in Huntington’s disease. ProgNeuropsychopharmacolBiol Psychiatry 27:255–265.
- Kwok JB (2010) Role of epigenetics in Alzheimer's and Parkinson's disease. Epigenomics 2:671-682.
- Wang YP, Song JF, Rao ZR, Mou DL (2004) Spinal cord neuronal apoptosis induced by triorthocresyl phosphate poisoning in hens. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing ZaZhi 22:19–21.
- Caughlan A, Newhouse K, Namgung U, Xia ZG (2004) Chlorpyrifos induces apoptosis in rat cortical neurons that is regulated by a balance between p38 and ERK/JNK MAP kinases. ToxicolSci 78:125-134.
- Emerick GL, Peccinini RG, de Oliveira GH (2010) Organophosphorus-induced delayed neuropathy: a simple and efficient therapeutic strategy ToxicolLett 192:238–244.
- Kusakawa G, Saito T, Onuki R, Ishiguro K, Kishimoto T, Hisanaga S (2000) Calpaindependentproteolytic cleavage of the p35 cyclin-dependent kinase 5 activator to p25. J. Biol. Chem 275: 17166–17172.
- Ranjbar A, Ghahremani MH, Sharifzadeh M, Golestani A, Ghazi-Khansari M, Baeeri M, Abdollahi M (2010) Protection by pentoxifylline of malathion-induced toxic stress and mitochondrial damage in rat brain. Hum ExpToxicol 29:851-864.
- Fortunato JJ, Feier G, Vitali AM, Petronilho FC, Dal-Pizzol F, Quevedo J (2006) Malathion induced oxidative stress in rat brain regions. Neurochem Res 31:671-678.
- Jain S, Banerjee BD, Ahmed RS, Arora VK, Mediratta PK (2013) Possible role of oxidative stress and brain derived neurotrophic factor in triazophos induced cognitive impairment in rats. Neurochem Res 38:2136-2147.
- Kim YH, Kim EY, Gwag BJ, Sohn S, Koh JY (1999) Zinc induced cortical neuronal death with features of apoptosis and necrosis: mediation by free radicals. Neuroscience 89:175–182.
- Russo A, Borrelli F, Campisi A, Acquaviva R, Raciti G, Vanella A (2003) Nitric oxide-related toxicity in cultured astrocytes: effect of Bacopamonniera. Life Sci 73:1517-1526.
- Venkatesh S, Ramachandran A, Zachariah A, Oommen A (2009) Mitochondrial ATP synthase inhibition and nitric oxide are involved in muscle weakness that occurs in acute exposure of rats to monocrotophos. ToxicolMech Methods 19:239-245.
- Akiyama H, Barger S, Barnum S, Bradt B, Bauer J, Cole GM, et al (2000) Inflammation and Alzheimer’s disease. Neurobiol Aging 21: 383-421.
- Liu L, Chan C (2014) The role of inflammasome in Alzheimer’s disease. Ageing Res. Rev 15: 6–15.
- Wenk GL, McGann K, Hauss-Wegrzyniak B, Rosi S (2003) The toxicity of tumor necrosis factor-alpha upon cholinergic neurons within the nucleus basalis and the role of norepinephrine in the regulation of inflammation: implications for Alzheimer’s disease. Neuroscience 121:719–729.
- Peress NS, Fleit HB, Perillo E, Kuljis R, Pezzullo C (1993) Identification of Fc gamma RI, II and III on normal human brain ramified microglia and on microglia in senile plaques in Alzheimer’s disease. J Neuroimmunol 48:71–79.
- Cacquevel M, Lebeurrier N, Cheenne S, Vivien D (2004) Cytokinesinneuroinflammation and Alzheimer’s disease. Curr. Drug Targets 5: 529-534.
- Arend WP, Palmer G, Gabay C (2008) IL−1, IL−18, and IL−33 families of cytokines. Immunol. Rev 223: 20-38.
- Chakraborty S, Kaushik DK, Gupta M, Basu A (2010) Inflammasome signaling at the heart of central nervous system pathology. J. Neurosci. Res 88: 1615-1631.
- Qiao X, Cummins DJ, Paul SM (2001) Neuroinflammation induced acceleration of amyloid deposition in the APPV717F transgenic mouse. Eur J Neurosci 14:474–482.
- Sunkaria A, Wani WY, Sharma DR, Gill KD (2012) Dichlorvos exposure results in activation induced apoptotic cell death in primary rat microglia. Chem Res Toxicol 25:1762-1770.
- Mawhinney L J, deRiveroVaccari JP, Dale GA, Keane RW, Bramlett HM (2011) Heightened inflammasome activation is linked to age related cognitive impairment in Fischer 344 rats. BMC Neurosci 12:123.
- Tan MS, Yu JT, Jiang T, Zhu XC, Tan L (2013) The NLRP3 inflammasome in Alzheimer’s disease. Mol. Neurobiol 48: 875–882.
- Coppieters N, Dragunow M (2011) Epigenetics in Alzheimer's disease: a focus on DNA modifications. Curr Pharm Des 17:3398-3412.
- Sanchez-Santed F, Colomina MT, Herrero Hernandez E (2015) Organophosphate pesticide exposure and neurodegeneration.
- Lein PJ, Bonner MR, Farahat FM, Olson JR, Rohlman DS, et al (2012) Experimental strategy for translational studies of organophosphorus pesticide neurotoxicity based on real world occupational exposures to chlorpyrifos. Neurotoxicology 33:660-668.
- Zaja-Milatovic S, Gupta RC, Aschner M, Milatovic D (2009) Protection of DFP-induced oxidative damage and neurodegeneration by antioxidants and NMDA receptor antagonist. ToxicolApplPharmacol 15:124-131.
- Liu X, Kim C, Yang J, Jemmerson R, Wang X (1996) Induction of apoptotic program in cellfree extracts: requirement for dATP and cytochrome c. Cell 86:147–157.
- Farooq RK, Isingrini E, Tanti A, LeGuisquet AM, Arlicot N, Minier F, et al (2012) Is unpredictable chronic mild stress (UCMS) are liable model to study depression induced neuroinflammation? Behav. Brain Res 231: 130–137.
- Fenske RA, Lu C, Barr D, Needham L (2002) Children’s exposure to chlorpyrifos and parathion in an agricultural community in central Washington State. Environ Health Perspect 110: 549–553.
- Yadav RS, Kumar N (2014) Lipid integration in neurodegeneration: An Overview of Alzheimer’s Disease. MolNeurobiol 50: 168-176.

