Keywords
Liver Diseases, Alcoholic; Pancreatitis, Chronic; Polymorphism, Genetic; Tumor Necrosis Factors
Abbreviations
kb: kilobases; LST1: leukocyte specific transcript 1; PBMC: peripheral blood mononuclear cell
INTRODUCTION
No single theory fully explains the clinical and morphological evolution of chronic pancreatitis. Theories to explain the pathogenesis of chronic pancreatitis include pancreatic duct obstruction by protein plugs [1], the toxic effects of ethanol and its metabolites [2], mounting oxidant stress [3] and recurrent acute pancreatitis [4]. More recent studies, which demonstrate the presence of activated cytotoxic T-cells within chronic pancreatitis resection specimens, suggest a primary pathogenic role for the immune system [5]. Similarly, a number of hypotheses regarding the pathogenesis of alcoholic liver disease exist but the role of immune responses leading to tissue injury has recently been emphasized. Studies have demonstrated circulating antibodies to acetaldehyde modified self-proteins, increased expression of proinflammatory cytokines and decreased anti-inflammatory cytokines. In particular, tumor necrosis factor (TNF) has emerged as the main candidate accounting for liver toxicity [6, 7, 8].
Alcohol is the major etiological agent for both chronic pancreatitis and alcoholic liver disease. However, as only approximately 10- 35% of all alcoholics develop alcoholic liver disease, 5% chronic pancreatitis, and only 1% develop both conditions [9, 10], there are clearly genetic or environmental cofactors that determine individual susceptibility to these diseases. A role for heredity in the pathogenesis of pancreatitis is suggested by familial clustering of alcoholic pancreatitis [11, 12] and by studies of genetic markers. HLA antigens [13], mutations of the cystic fibrosis transmembrane conductance regulator gene [14, 15] and SPINK1 gene mutations [16] have been associated with susceptibility to alcoholic chronic pancreatitis. For alcoholic liver disease, twin studies [17] and associations with polymorphisms of the genes encoding the cytochrome P-450-2E1 [18, 19], alcohol dehydrogenase [20], TNF [21] and interleukin-10 [22] also suggest a genetic component. It is likely that the expression of both these disease phenotypes, in common with other complex human diseases, is influenced by strong environmental risk factors plus the possession of a range of susceptibility genes.
TNF is produced predominantly by macrophage/monocytes but also by T and Blymphocytes neutrophils and endothelial cells. Its functions include a pivotal role in the initiation of the release of other proinflammatory mediators, activation of endothelial cells, upregulation of the expression of intercellular adhesion molecules, induction of T and B-lymphocyte proliferation, enhancement of T-cell mediated cytotoxicity and increased MHC (class I and class II) expression [23, 24]. Stable interindividual variations exist in levels of production of TNF, suggesting inherited individual differences [25]. Family studies show that up to 60% of the variability in TNF production between individuals may be genetically determined [26]. Association studies have been widely used to investigate the putative association of TNF polymorphisms and susceptibility to various diseases. TNF polymorphisms have been associated with susceptibility and outcome of various infectious [27, 28, 29], inflammatory [30, 31] and neoplastic [32, 33] diseases.
In acute pancreatitis, the haplotype TNFa2b1c2 has a reduced frequency among those of alcoholic aetiology [34]. The finding of this genetic difference among alcoholics who develop acute pancreatitis led us to hypothesize that this locus may play an even more important role in the development of chronic pancreatitis and liver disease, where alcohol predominates as an aetiological factor.
MATERIALS AND METHODS
Study Subjects and Diagnostic Criteria
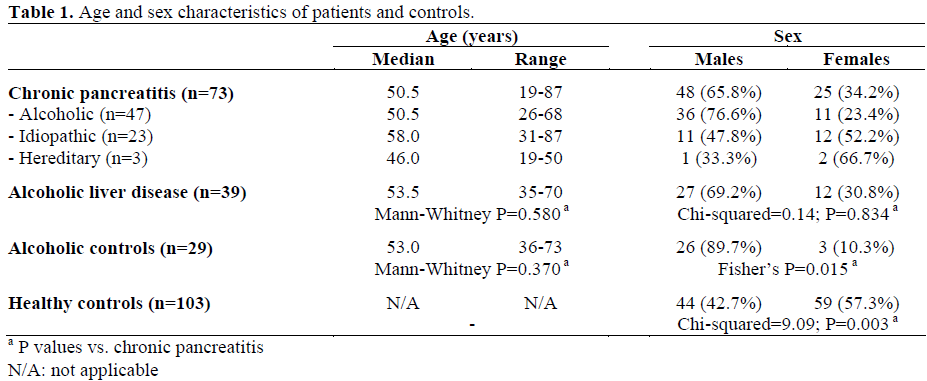
DNA was extracted from 73 patients with chronic pancreatitis (47 alcoholic, 23 idiopathic and 3 hereditary), 103 healthy controls, 39 patients with alcoholic liver disease and 29 alcoholics with no clinical evidence of either liver or pancreatic disease. Demographic data are provided in Table 1. Analysis of the demographic data showed no significant difference with respect to age but significant differences were seen for male/female sex ratios between chronic pancreatitis and alcoholic controls (Fisher’s P=0.015) and between chronic pancreatitis and healthy controls (chi-squared=9.09; P=0.003). All patients and controls were Caucasoid.
The diagnostic criteria for chronic pancreatitis was based upon the classification system adopted at the Zurich Workshop of 1996 [35]. For diagnosis, a typical history of recurrent clinical acute pancreatitis was required in addition to one or more of the following: pancreatic calcification, ductal lesions [36], steatorrhoea reduced by enzyme supplementation, and histology. The definition of “definite alcoholic chronic pancreatitis” was based upon the above plus an excessive alcohol intake (greater than 80 g/day for more than 2 years) [35].
Alcoholic liver disease patients were patients with acute alcoholic hepatitis or alcohol alcoholinduced cirrhosis who were without clinical evidence of pancreatitis, attending the Gastroenterology Department, Derriford Hospital, Plymouth, UK. All had drunk at least 8 units of alcohol per day for at least two years.
Alcoholic controls were patients undergoing treatment for alcoholism at the Broadreach Rehabilitation Unit, Plymouth, UK. All had drunk at least 56 units of alcohol per week for two or more years. They were without clinical or biochemical evidence of either chronic pancreatitis or alcoholic liver disease.
Healthy controls were Caucasoid cord-blood samples following a normal, healthy obstetric delivery at Derriford Hospital, Plymouth, UK.
DNA Preparation
High molecular weight DNA was prepared from peripheral venous blood using the Nucleon II DNA extraction kit (Scotlab, Lanarkshire, UK).
TNF Microsatellite Typing
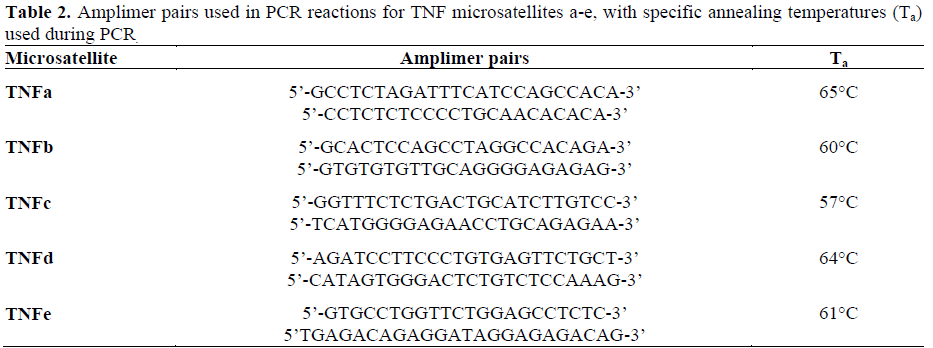
The five microsatellite loci: TNFa, TNFb, TNFc, TNFd, TNFe were amplified by the polymerase chain reaction as a one-step procedure using oligonucleotide amplimers (MWG-Biotech, Buckinghamshire, UK) synthesized from previously described sequences [37] (Table 2). One-hundred to 500 ng of genomic DNA was added to a final reaction volume of 20 mL with 0.5 mM of each amplimer pair (reverse amplimer 5’; end labeled with 50,000 cpm of 32P), 10 mM Tris- HCl (pH 9.0), 50 mM KCl, 2.5 mM MgCl2, 0.1% Triton X-100, 300 mM of each deoxyribonucleoside 5'-triphosphates (dNTPs), and 0.8 units of Taq DNA polymerase (HT Biotechnology, Cambridge, UK). PCR was performed in a cyclogene thermocycler (Techne, Cambridge, UK) under the following conditions: 95°C for 4 minutes, then 30 cycles of 95°C for 2 minutes, 2 minutes at the specific annealing temperature (Table 2), 62°C for 2 minutes, followed by an extension period of 10 minutes at 68°C. Nonradioactive PCR products underwent electrophoresis on 1% agarose gel, to confirm the presence of product of anticipated size. Amplification products (6 mL) were then mixed with 3 mL of formamide containing Stop solution (Amersham Life Science, Buckinghamshire, UK) and then separated on a 6% polyacrylamide gel containing 8 M urea at 1,700 V for 2.5 hours (TNFa, TNFc, TNFd, TNFe) or 3 hours (TNFb). After drying, the gels were exposed to Kodak XLS5 X-ray film (Scientific Imaging Systems, Cambridge, UK) with intensifying screens at -80°C for 18 hours.
Haplotype Designation
Alleles that combine in non-random association across the TNF five locus microsatellite system are termed haplotypes. Haplotype studies are more powerful than gene frequency studies [38]; therefore TNF haplotype data have been presented rather than a comparison of allele frequencies. These TNF haplotypes have previously been extensively characterized, by maximum likelihood estimates in population and family studies and their existence confirmed in homozygous cell lines [39, 40]. Associations between TNF-alpha and TNF-beta secretion levels and the frequent Caucasian TNF five locus microsatellite haplotypes have been determined in macrophages and lymphoid cells in response to a variety of stimuli, including lipopolysaccharide and T-cell mitogens [41]. TNF alleles at each of the fivemicrosatellite loci were ascribed to a haplotype, as previously defined [39, 40, 41], for each sample. Table 3 provides the definition of each haplotype and their secretory status, under specified conditions.

ETHICS
Local Research Ethics Committee approval was obtained. In accordance with the American Society of Human Genetics (ASHG) guidelines on genetic research [42], informed consent was obtained from all subjects in whom specimens were obtained prospectively, i.e. those with pancreatitis and those with alcoholic liver disease. Additional clinical, pathological and demographic information was obtained before subject identifiers were removed and the samples “anonymised”. The healthy control study utilized retrospective samples collected anonymously, in whom there was no possibility, or need, to obtain consent.
STATISTICS
Comparison of haplotype frequencies between patients and controls were made in 2x2 contingency tables using the chi-squared method. Corrections were made, if necessary, for small sample numbers using Fisher’s exact test (i.e. where a cell in a 2x2 contingency table contained a number less than 5). The odds ratio (OR) and the 95% confidence interval (CI) were also evaluated. Because a single hypothesis was being tested, i.e. that our previously reported TNF haplotype association with acute alcoholic pancreatitis [34], would show a similar association with chronic pancreatitis, a two tailed P value of less than 0.05 was considered statistically significant, without further correction [43]. A recent review of immunogenetics has confirmed the legitimacy of this approach and states: “the alternative to correction is to verify (replicate) in subsequent studies” [44]. Ages were compared by means of the Mann-Whitney U test. The SPSS 8.0 for Windows was used to analyse the data.
RESULTS
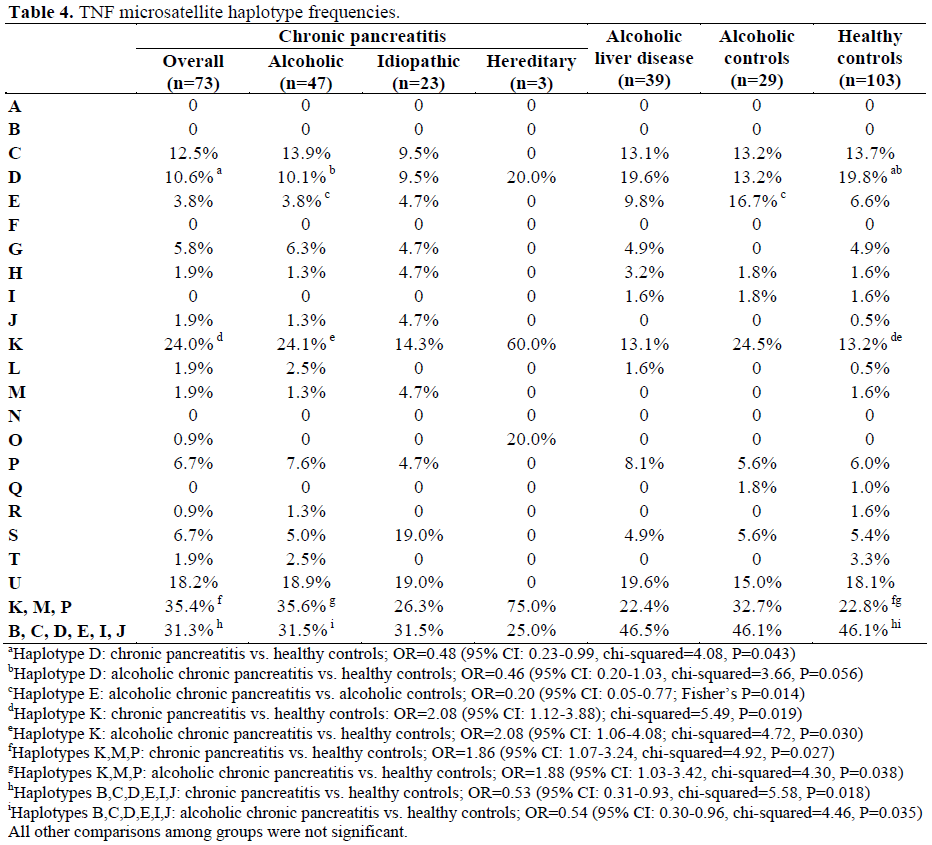
The TNF a6b5c1d3e3 Haplotype Is Positively Associated with Chronic Pancreatitis; the TNF a2b3c1d1e3 and a2b5c2d4e3 Haplotypes are Negatively Associated (Table 4)
The intermediate/low TNF secreting haplotype a6b5c1d3e3 (K) was overrepresented in patients with chronic pancreatitis compared to healthy controls. It was present in 25 out of 104 (24.0%) identified haplotypes in patients with chronic pancreatitis compared with 24 out of 182 (13.2%) identified haplotypes among healthy controls. The OR was 2.08 (95% CI: 1.12- 3.88; chi-squared=5.49, P=0.019). This haplotype was specifically over-represented in patients with alcoholic chronic pancreatitis compared to healthy controls. It was present in 19 out of 79 (24.1%) identified haplotypes in patients with alcoholic chronic pancreatitis compared with 24 out of 182 (13.2%) haplotypes among healthy controls. The OR was 2.08 (95% CI: 1.06-4.08; chisquared= 4.72, P=0.030).
The high TNF secreting haplotype a2b3c1d1e3 (D) was under-represented in patients with chronic pancreatitis compared to healthy controls. It was present in 11 out of 104 (10.6%) identified haplotypes in patients with chronic pancreatitis compared with 36 out of 182 (19.8%) haplotypes among healthy controls. The OR was 0.48 (95% CI: 0.23- 0.99; chi-squared=4.08, P=0.043). This haplotype was specifically under-represented in patients with alcoholic chronic pancreatitis compared to healthy controls. It was present in 8 out of 79 (10.1%) identified haplotypes in patients with alcoholic chronic pancreatitis compared with 36 out of 182 (19.8%) haplotypes among healthy controls. The OR was 0.46 (95% CI: 0.20-1.03; chisquared= 3.66, P=0.056). Furthermore, a similar high TNF secreting haplotype a2b5c2d4e3 (E) was underrepresented in patients with alcoholic chronic pancreatitis compared to alcoholic controls. It was present in 3 out of 79 (3.8%) identified haplotypes in patients with alcoholic chronic pancreatitis compared with 9 out of 54 (16.7%) haplotypes among alcoholic controls. The OR was 0.20 (95% CI: 0.05-0.77; Fisher’s P=0.014).
Low-Intermediate TNF Secreting Haplotypes Are Positively Associated with Chronic Pancreatitis; High Secreting Haplotypes Are Negatively Associated (Table 4)
The low-intermediate secreting TNF haplotypes (K, M, P) were over-represented in patients with chronic pancreatitis compared to healthy controls. They were present in 34 out of 96 (35.4%) identified haplotypes in patients with chronic pancreatitis compared to 38 out of 167 (22.8%) haplotypes in healthy controls. The OR was 1.86 (95% CI: 1.07- 3.24; chi-squared=4.92, P=0.027). These haplotypes were specifically over-represented in patients with alcoholic chronic pancreatitis compared to healthy controls. They were present in 26 out of 73 (35.6%) identified haplotypes in patients with alcoholic chronic pancreatitis compared to 38 out of 167 (22.8%) haplotypes in healthy controls. The OR was 1.88 (95% CI: 1.03-3.42; chisquared= 4.30, P=0.038). All high secreting TNF haplotypes (B, C, D, E, I, J) were under-represented in patients with chronic pancreatitis compared to healthy controls. They were present in 30 out of 96 (31.3%) identified haplotypes in patients with chronic pancreatitis compared to 77 out of 167 (46.1%) haplotypes in healthy controls. The OR was 0.53 (95% CI: 0.31-0.93; chisquared= 5.58, P=0.018). These haplotypes were specifically under-represented in patients with alcoholic chronic pancreatitis compared to healthy controls. They were present in 23 out of 73 (31.5%) identified haplotypes in patients with alcoholic chronic pancreatitis compared to 77 out of 167 (46.1%) haplotypes in healthy controls. The OR was 0.54 (95% CI: 0.30-0.96; chisquared= 4.46, P=0.035).
No Association between Idiopathic Chronic Pancreatitis or Alcoholic Liver Disease and TNF Haplotypes (Table 4)
Trends towards association of the alcoholic liver disease patients with high secreting microsatellite haplotypes were found, e.g. for haplotype D (a2b3c1d1e3) when compared to alcoholic pancreatitis, but these did not reach statistically significant levels (chisquared= 2.56, P=0.100). No significant association between idiopathic chronic pancreatitis and TNF haplotypes was observed.
DISCUSSION
The TNF gene cluster is located within the central MHC (class III region) on the short arm of chromosome six. It spans 12 kilobases (kb) and maps 250 kb centromeric to the HLA-B locus and 850 kb to the class II region [45]. The locus contains genes coding for TNF, lymphotoxin alpha (previously TNFbeta), lymphotoxin beta and leukocyte specific transcript 1 (LST1) [46]. Two clusters of single nucleotide polymorphisms and five microsatellite markers are also found within this locus [47]. Stable interindividual variations exist in levels of production of TNF, suggesting inherited individual differences [25]. Family studies show that up to 60% of the variability in TNF production between individuals may be genetically determined [26] and this has been correlated with specific MHC alleles and ancestral haplotypes [48]. An important feature of the MHC is the strong linkage disequilibrium between particular alleles across this region. This has prompted speculation that the association between high TNF production and HLA-DR alleles may be related to polymorphism within the TNF region itself. Allele 2 of the TNF promoter polymorphism at position -308 has been correlated with high TNF production by peripheral blood mononuclear cells [49] and a seven-fold increase of TNF transcription in a CAT reporter gene transiently transfected into a human B cell line [50]. The highly polymorphic TNF microsatellite markers may be more informative, with high levels of TNF production associated with TNFa2, TNFc2 and TNFd3 alleles and low levels with TNFa6 [51, 52]. Thus, the TNF response correlates with a number of genetic markers all mapped within a region with strong linkage disequilibrium.
Linkage analysis has been used successfully to find major gene effects but has limited power to detect more modest effects. Association studies, which utilize candidate genes, have far greater power to delineate the genetics of complex human diseases [53]. This approach has been widely used in relation to the putative association of TNF polymorphisms and susceptibility to various diseases. In Crohn’s disease, the five microsatellite haplotype TNFa2b1c2d4e1 is overrepresented compared to patients with ulcerative colitis and to healthy controls [30]. This haplotype is in linkage disequilibrium with HLA-DR1/DQ5. A different microsatellite haplotype, TNFa6b5c1d3e, has been described in rheumatoid arthritis [31]. In contrast to Crohn’s disease, this association is independent of HLA linkage. TNF polymorphisms have also been associated with susceptibility and outcome of various infectious [27, 28, 29] and neoplastic [32, 33] diseases.
In a previous study, we evaluated the association of the TNF and IL-10 genes with acute pancreatitis and resultant disease severity [34]. When patients were stratified according to disease aetiology, the haplotype TNFa2b1c2 showed a trend toward reduced frequency in the group with alcohol induced acute pancreatitis. TNFa2b1c2 was present in 4.8% of individuals with alcohol induced acute pancreatitis compared with 24.3% of the controls, 31.9% of patients with non-alcoholic pancreatitis. However, this was not significant after correction for multiple comparisons. The finding of a genetic difference among alcoholics who develop acute pancreatitis in our original study suggested that this locus may play an even more important role in the development of chronic pancreatitis, where alcohol plays an even more predominant role as an etiological factor. Therefore, we expanded our study to determine the frequencies of TNF gene markers in a larger group of patients with alcohol induced chronic pancreatitis. The entry criteria for the current study differs from our previous study. For the original study of patients with acute pancreatitis a consistent clinical history, appropriate radiological evidence, and a serum amylase level greater than three times the upper limit of normal, were required. Patients were then classified as having mild disease or severe local or systemic disease according to criteria defined by the Atlanta convention. The entry criteria for the current study of chronic pancreatitis was based upon the classification system adopted at the Zurich Workshop, 1996 [35] i.e. a typical history of recurrent clinical acute pancreatitis was required in addition to one or more of the following: pancreatic calcification, ductal lesions [36], steatorrhoea reduced by enzyme supplementation and histology. The finding that the a6b5c1d3e3 is positively associated with chronic pancreatitis and the haplotypes a2b5c2d1e3 and a2b5c2d1e3 are negatively associated, confirms the association between alcoholic pancreatitis and genetic markers of reduced TNF production.
Other studies of the TNF gene and its putative association with chronic pancreatitis include a recent investigation that demonstrated that the frequencies of the TNF promoter polymorphisms at position -238 and -308 in patients with alcoholic chronic pancreatitis, idiopathic pancreatitis and those with the N34S mutation of the SPINK 1 gene, did not differ significantly from the control group. However, the variant TNF-238A was a risk factor for disease manifestation in families with hereditary pancreatitis [54].
Histological examination of the normal pancreas reveals that inflammatory cells are a rare finding. In addition, MHC class I antigens, which are expressed by most nucleated somatic cells, are not present on exocrine pancreatic cells. Normal acinar and ductal pancreatic epithelial cells do not express MHC class II determinants [55, 56]. In chronic pancreatitis, this is radically altered. Lymphocytic infiltrates are common, with the cytotoxic CD8+ T-cell subset predominating [56, 57, 58]. Abnormal expression of class I or class II MHC determinants, or both, by exocrine epithelial cells was demonstrated in 89% of ninety-three cases of chronic pancreatitis [56]. Until recently, the cellular infiltrate was not considered to be of primary pathophysiological importance but merely an epiphenomenon consequent upon other primary pathological changes in the pancreas. Recent findings however, provide increasing evidence that immunologic effector mechanisms might be instrumental in the induction of chronic pancreatitis. Hunger has demonstrated that CD8+ T-cells are activated in cellular infiltrates of alcoholic chronic pancreatitis, using perforin mRNA as a specific in vivo activation marker [59]. In MHC class II deficient mice, an animal model originally designed to study ulcerative colitis, Vallance et al. unexpectedly observed the development of an immune-based pancreatitis with selective loss of exocrine cells and function. Furthermore, CD8+ cells alone were able to adoptively transfer the disease to athymic mice and, in their absence, the remaining cells were unable to induce any pancreatic pathology [60]. Autoreactive Tcells normally exist within the immune system but are kept quiescent, or anergic, by other regulatory T lymphocytes. In cases in which immune balance is disrupted, such as the immunodeficiency seen in the MHC class II-deficient mice, the loss of regulatory Tcells may allow self-reactive lymphocytes to attack autoantigens. Alteration in the distribution and function of peripheral blood lymphocytes in patients with chronic pancreatitis has been reported [61]. Gansauge et al. [61] found an increase of both CD4+ and CD8+ T-cells among chronic pancreatitis patients. Evidence of reduced immunosurveillance was detected upon stimulation with anti-CD3+ antibodies. Pancreatic head resection restored this suppressed immune function. Finally, demonstration of a direct relationship between the degree of perineural inflammation and pain strongly supports the hypothesis of neuroimmune interaction as an important, if not predominant, factor in pain generation in chronic pancreatitis [62]. TNF, a mediator that plays a pivotal role in the induction of T-cell proliferation, enhancement of T-cell mediated cytotoxicity and increased MHC (class I and class II) expression [23, 24], was therefore chosen for examination in our study.
The finding that a haplotype containing an allele associated with diminished TNF production (a6b5c1d3e3) is positively associated with chronic pancreatitis while haplotypes with alleles associated with enhanced TNF production (a2b5c2d1e3 and a2b5c2d4e3) are negatively correlated seems at odds with our understanding of the role of TNF in the pathogenesis of chronic pancreatitis. TNF, however, is the classic immunological double-edged sword and evidence exists that may explain why individuals who are genetically programmed to secrete relatively low amounts of TNF may be more susceptible to developing chronic pancreatitis.
Jacob and McDevitt have demonstrated that (NZBxNZW)F1 hybrid mice develop a severe autoimmune disease similar to systemic lupus erythematosus. These mice are deficient in TNF production and this correlates with a restriction fragment length polymorphism in the TNF gene. Furthermore, replacement therapy with recombinant TNF induced a significant delay in the development of disease [63]. The assumption that TNF is a culprit in the pathogenesis of immunemediated diseases has also been challenged in a phase 1 clinical study with a monoclonal anti-TNF antibody in multiple sclerosis [64]. Treatment caused immune activation and an increase in disease activity.
An explanation for a detrimental effect being observed with diminished TNF production in chronic pancreatitis may be provided by studies demonstrating that TNF can induce Tcell apoptosis in vivo [65, 66, 67]. Furthermore, it was CD8+ rather than CD4+ T-cells that were the principal targets of TNF action. It will be recalled that it was CD8+ cells that alone were capable of adoptively transferring an immune-based pancreatitis [60]. Alcoholics may be at greater risk of expressing the disease phenotype because alcohol downregulates TNF activity in peripheral blood mononuclear cells (PBMCs) still further [68]. Thus, it may be that a reduced capacity to produce TNF may be responsible for the induction of chronic pancreatitis, via a failure to delete autoreactive T-cells by apoptosis
A number of hypotheses exist regarding the pathogenesis of alcoholic liver disease [69]. However, TNF has emerged as a key mediator of the hepatic necro-inflammation that is associated with excessive alcohol intake. Increased TNF production by peripheral blood monocytes and Kupffer cells has been shown both in animal models and patients with alcoholic liver disease, correlating with liver function and mortality [70, 71, 72, 73]. Moreover, TNF is directly cytotoxic to sensitized hepatocytes [74] and was the major toxicity encountered when human recombinant TNF was administered therapeutically for cancer [75]. Finally, a significant excess of the TNF -238 G->A polymorphism, which is associated with increased TNF expression, is associated with susceptibility to alcoholic steatohepatitis [21]. In our study, trends towards the association of alcoholic liver disease with high secreting microsatellite haplotypes were found, e.g. for haplotype D (a2b3c1d1e3) when compared to alcoholic pancreatitis, but these did not reach statistically significant levels. If confirmed, this would be consistent with the view that TNF plays a genetically-determined, proinflammatory and cytotoxic role in alcoholic liver disease.
References
- Sarles H, Bernard JP, Johnson C. Pathogenesis and epidemiology of chronic pancreatitis. Annu Rev Med 1989; 40:453-68. [PMID 2658760]
- Bordalo O, Goncalves D, Noronha M, Cristina ML, Salgadinho A, Dreiling DA. Newer concept for the pathogenesis of chronic alcoholic pancreatitis. Am J Gastroenterol 1977; 68:278-285. [PMID 596358]
- Braganza JM. A framework for the aetiogenesis of chronic pancreatitis. Digestion 1998; 59(Suppl 4):1-12. [PMID 9832631]
- Kloppel G, Maillet B. Chronic pancreatitis: evolution of the disease. Hepatogastroenterology 1991; 38:408-12. [PMID 1765357]
- Hunger RE, Mueller C, Z'Graggen K, Friess H, Buchler MW. Cytotoxic cells are activated in cellular infiltrates of alcoholic chronic pancreatitis. Gastroenterology 1997; 112:1656-63. [PMID 9136845]
- Xu D, Thiele GM, Beckenhauer JL, Klassen LW, Sorrell MF, Tuma DJ. Detection of circulating antibodies to malondialdehyde-acetaldehyde adducts in ethanol-fed rats. Gastroenterology 1998; 115:686-92. [PMID 9721166]
- Fang C, Lindros KO, Badger TM, Ronis MJ, Ingelman-Sundberg M. Zonated expression of cytokines in rat liver: effect of chronic ethanol-induced toxicity and the cytochrome P450 2EI inhibitor chlormethiazole. Hepatology 1998; 27:1304-10. [PMID 9581684]
- Neuman MG, Shear NH, Bellentani S, Tiribelli C. Role of cytokines in ethanol-induced cytotoxicity in vitro in hep G2 cells. Gastroenterology 1998; 115:157- 66. [PMID 9649471]
- Gorelick FS, Robles-Diaz G. Alcohol and pancreatitis. In: Buchler MW, Friess H, Uhl W, Malfertheiner P, eds. Chronic Pancreatitis: Novel Concepts in Biology and Therapy. Oxford, United Kingdom: Blackwell Publishing, 85-100.
- Sorensen TI, Orholm M, Bentsen KD, Hoybye G, Eghoje K, Christoffersen P. Prospective evaluation of alcohol abuse and alcoholic liver injury in men as predictors of development of cirrhosis. Lancet 1984; 2:241-4. [PMID 6146805]
- Sarles H, Sarles JC, Camatte R, Muratore R, Gaini M, Guien C, et al. Observations on 205 confirmed cases of acute pancreatitis, recurring pancreatitis and chronic pancreatitis. Gut 1965; 6:545-9. [PMID 5857891]
- Marks IN, Bank S, Louw JH. Chronic pancreatitis in the Western Cape. Digestion 1973; 9:447-53. [PMID 4784706]
- Wilson JS, Gossat D, Tait A, Rouse S, Juan XJ, Pirola RC. Evidence for an inherited predisposition to alcoholic pancreatitis. A controlled HLA typing study. Dig Dis Sci 1984; 29:727-30. [PMID 6589150]
- Sharer N, Schwarz M, Malone G, Howarth A, Painter J, Super M, et al. Mutations of the cystic fibrosis gene in patients with chronic pancreatitis. N Engl J Med 1998; 339:645-52. [PMID 9725921]
- Cohn JA, Friedman KJ, Noone PG, Knowles MR, Silverman LM, Jowell PS. Relation between mutations of the cystic fibrosis gene and idiopathic pancreatitis. N Engl J Med 1998; 339:653-8. [PMID 9725922]
- Witt H, Luck W, Becker M, Bohmig M, Kage A, Truninger K, et al. Mutation in the SPINK1 trypsin inhibitor gene, alcohol use, and chronic pancreatitis. JAMA 2001; 285:2716-7. [PMID 11386926]
- Hrubec Z, Omenn GS. Evidence of genetic predisposition to alcoholic cirrhosis and psychosis: twin concordances for alcoholism and its biological end points by zygosity among male veterans. Alcohol ClinExp Res 1981; 9:306-9. [PMID 7018299]
- Grove J, Brown AS, Daly AK, Bassendine MF, James OF, Day CP. The RsaI polymorphism of CYP2E1 and susceptibility to alcoholic liver disease in Caucasians: effect on age of presentation and dependence on alcohol dehydrogenase genotype. Pharmacogenetics1998 ;8:335-42. [PMID 9731720]
- Tsutsumi M, Takada A, Wang JS. Genetic polymorphisms of cytochrome P4502E1 related to the development of alcoholic liver disease. Gastroenterology 1994; 107:1430-5. [PMID 7926507]
- Frenzer A, Butler WJ, Norton ID, Wilson JS, Apte MV, Pirola RC, et al. Polymorphism in alcoholmetabolizing enzymes, glutathione S-transferases and apolipoprotein E and susceptibility to alcohol-induced cirrhosis and chronic pancreatitis. J GastroenterolHepatol 2002; 17:177-82. [PMID 11966948]
- Grove J, Daly AK, Bassendine MF, Day CP. Association of a Tumor Necrosis Factor promoter polymorphism with susceptibility to alcoholic steatohepatitis. Hepatology 1997; 26:143-6. [PMID 9214463]
- Grove J, Daly AK, Bassendine MF, Gilvarry E, Day CP. Interleukin 10 promoter region polymorphisms and susceptibility to advanced alcoholic liver disease. Gut 2000; 46:540-5. [PMID 10716685]
- Cohen MC, Cohen S. Cytokine function: a study in biologic diversity. Am J ClinPathol 1996; 105:589-98. [PMID 8623768]
- Papadakis KA, Targan SR. Tumor Necrosis Factor: biology and therapeutic inhibitors. Gastroenterology 2000; 119:1148-57. [PMID 11040201]
- Molvig J, Baek L, Christensen P, Manogue KR, Vlassara H, Platz P, et al. Endotoxin stimulated human monocyte secretion of interleukin 1, tumor necrosis factor alpha and prostaglandin E2 shows stable interindividual differences. Scand J Immunol 1988; 27:705-16. [PMID 3260683]
- Westendorp RG, Langermans JA, Huizinga TW, Elouali AH, Verweij CL, Boomsma DI, Vandenbroucke JP. Genetic influences on cytokine production and fatal meningococcal disease. Lancet 1997; 349:170-3. [PMID 9111542]
- Cabrera M, Shaw MA, Sharples C, Williams H, Castes M, Convit J, et al. Polymorphism in tumor necrosis factor genes associated with mucocutaneous leishmaniasis. J Exp Med 1995; 182:1259-64. [PMID 7595196]
- Mira JP, Cariou A, Grall F, Delclaux C, Losser MR, Heshmati F, et al. Association of TNF2, a TNFalpha promoter polymorphism, with septic shock susceptibility and mortality. JAMA 1999; 282:561-8. [PMID 10450718]
- McGuire W, Hill AV, Allsopp CE, Greenwood BM, Kwiatkowski D. Variation in the TNF-alpha promoter region associated with susceptibility to cerebral malaria. Nature 1994; 371:508-11. [PMID 7935762]
- Plevy SE, Targan SR, Yang H, Fernandez D, Rotter JI, Toyoda H. Tumor necrosis factor microsatellites define a Crohn’s disease-associated haplotype on chromosome 6. Gastroenterology 1996; 110:1053-60. [PMID 8612993]
- Mulcahy B, Waldron-Lynch F, McDermott MF, Adams C, Amos CI, Zhu DK, et al. Genetic variability in the Tumor Necrosis Factor-Lymphotoxin region influences susceptibility to rheumatoid arthritis. Am J Hum Genet 1996; 59:676-83. [PMID 8751869]
- Chouchane L, Ahmed SB, Baccouche S, Remadi S. Polymorphism in the tumor necrosis factor-alpha promoter region and in the heat shock protein 70 genes associated with malignant tumors. Cancer 1997; 80:1489-96. [PMID 9338474]
- Gallagher G, Lindemann M, Oh HH, Ferencik S, Walz MK, Schmitz A, et al. Association of the TNFa2 microsatellite allele with the presence of colorectal cancer. Tissue Antigens 1997; 50:47-51. [PMID 9243755]
- Sargen K, Demaine AG, Kingsnorth AN. Cytokine gene polymorphisms in acute pancreatitis. JOP. J Pancreas (Online) 2000; 1:24-35. [PMID 11852287]
- Ammann RW. A clinically based classification system for alcoholic chronic pancreatitis: summary of an international workshop on chronic pancreatitis. Pancreas 1997; 14:215-21. [PMID 9094150]
- Axon AT, Classen M, Cotton PB, Cremer M, Freeny PC, Lees WR. Pancreatography in chronic pancreatitis: international definitions. Gut 1984; 25:1107-12. [PMID 6479687]
- Udalova IA, Nedospasov SA, Webb GC, Chaplin DD, Turetskaya RL. Highly informative typing of the human TNF locus using six adjacent polymorphic markers. Genomics 1993;16:180-6. [PMID 8486354]
- Crouau-Roy B, Briant L, Bouissou C, Stravropoulos C, Pociot F, Cambon-Thomsen A, Clayton J. Tumor necrosis factor microsatellites in four European populations. Hum Immunol 1993; 38:213-6. [PMID 8106279]
- Jongeneel CV, Briant L, Udalova IA, Sevin A, Nedospasov SA, Cambon-Thomsen A. Extensive genetic polymorphism in the human tumor necrosis factor region and relation to extended HLA haplotypes. ProcNatlAcadSci 1991; 88:9717-21. [PMID 1946393]
- Nedospasov SA, Udalova IA, Kuprash DV, Turetskaya RL. DNA sequence polymorphism at the human tumor necrosis factor (TNF) locus. Numerous TNF/lymphotoxin alleles tagged by two closely linked microsatellites in the upstream region of thelymphotoxin (TNF-beta) gene. J Immunol 1991; 147:1053-9. [PMID 1861069]
- Weissensteiner T, Lanchbury JS. TNFB polymorphisms characterize three lineages of TNF region microsatellite haplotypes. Immunogenetics 1997; 47:6-16. [PMID 9382915]
- ASHG report. Statement on informed consent for genetic research. Am J Hum Genet 1996; 59:471-4. [PMID 8755936]
- Perneger TV. What’s wrong with Bonferroni adjustments. BMJ 1998; 316:1236-8. [PMID 9553006]
- Donaldson PT. Genetics of liver disease: Immunogenetics and disease pathogenesis. Gut 2004; 53:599-608. [PMID 15016758]
- Makhatadze NJ. Tumor necrosis factor locus: Genetic organization and biological implications. Hum Immunol 1998; 59:571-9. [PMID 9757913]
- Holzinger I, de Baey A, Messer G, Kick G, Zwierzina H, Weiss EH. Cloning and genomic characterization of LST1: a new gene in the human TNF region. Immunogenetics 1995; 42:315-22. [PMID 7590964]
- Knight JC, Kwiatkowski D. Inherited variability of tumor necrosis factor production and susceptibility to infectious disease. ProcAssoc Am Physicians 1999; 111:290-8. [PMID 10417736]
- Jacob CO, Fronek Z, Lewis GD, Koo M, Hansen JA, McDevitt HO. Heritable Major histocompatibility complex class II associated differences in production of tumor necrosis factor alpha: Relevance to genetic predisposition to systemic lupus erythematosus. ProcNatlAcadSci USA 1990; 87:1233-7. [PMID 2105500]
- Bouma G, Crusius JB, Oudkerk Pool M, Kolkman JJ, von Blomberg BM, Kostense PJ, et al. Secretion of tumor necrosis factor alpha and lymphotoxin alpha in relation to polymorphisms in the TNF genes and HLADR alleles. Relevance for inflammatory bowel disease. Scand J Immunol 1996: 43:456-63. [PMID 8668926]
- Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. ProcNatlAcadSci USA 1997; 94:3195-9. [PMID 9096369]
- Pociot F, Briant L, Jongeneel CV, Molvig J, Worsaae H, Abbal M, et al. Association of tumor necrosis factor (TNF) and class II major histocompatibility complex alleles with the secretion of TNF-alpha and TNF-beta by human mononuclear cells: a possible link to insulin dependent diabetes mellitus. Eur J Immunol 1993; 23:224-31. [PMID 8093442]
- Turner DM, Grant SC, Lamb WR, Brenchley PE, Dyer PA, Sinnott PJ, Hutchinson IV. A genetic marker of high TNF-alfa production in heart transplantrecipients. Transplantation 1995; 60:1113-7. [PMID 7482718]
- Risch N, Merikangas K. The future of genetic studies of complex human diseases. Science 1996; 273:1516-7. [PMID 8801636]
- Beranek H, Teich N, Witt H, Schulz HU, Mossner J, Keim V. Analysis of tumour necrosis factor alpha and interleukin 10 promoter variants in patients with chronic pancreatitis. Eur J GastroenterolHepatol 2003; 15:1223-7. [PMID 14560157]
- Bedossa P, Bacci, Lemaigre G, Martin E. Lymphocyte subsets and HLA-DR expression in normal pancreas and chronic pancreatitis. Pancreas 1990; 5:415-20. [PMID 1974351]
- Jalleh RP, Gilbertson JA, Williamson RC, Slater SD, Foster CS. Expression of major histocompatibility antigens in human chronic pancreatitis. Gut 1993; 34:1452-7. [PMID 8244120]
- Bockman DE, Buchler M, Malfertheiner P, Beger HG. Analysis of nerves in chronic pancreatitis. Gastroenterology 1988; 94:1459-69. [PMID 3360267]
- Emmrich J, Weber I, Nausch M, Sparmann G, Koch K, Seyfarth M, et al. Immunohistochemical characterization of the pancreatic cellular infiltrate in normal pancreas, chronic pancreatitis and pancreatic carcinoma. Digestion 1998; 59:192-8. [PMID 9643678]
- Hunger RE, Mueller C, Z’Graggen K, Friess H, Buchler M. Cytotoxic cells are activated in cellular infiltrates of alcoholic chronic pancreatitis. Gastroenterology 1997; 112:1656-63. [PMID 9136845]
- Vallance BA, Hewlett BR, Snider D, Collins SM. T-cell mediated exocrine pancreatic damage in major histocompatibility complex class II-deficient mice. Gastroenterology 1998; 115:978-87. [PMID 9753501]
- Gansauge F, Gansauge S, Eh M, Schlosser W, Ramadani M, Kern P, et al. Distributional and functional alterations of immunocompetent peripheral blood lymphocytes in patients with chronic pancreatitis. Ann Surg 2001; 233:365-70. [PMID 11224624]
- Di Sebastiano, di Mola FF, Bockman DE, Friess H, Buchler MW. Chronic pancreatitis: the perspective of pain generation by neuroimmune interaction. Gut 2003; 52:907-11. [PMID 12740353]
- Jacob CO, McDevitt HO. Tumor necrosis factoralpha in murine autoimmune 'lupus' nephritis. Nature 1988; 331:356-8. [PMID 2893286]
- van Oosten BW, Barkhof F, Truyen L, Boringa JB, Bertelsmann FW, von Blomberg BM, et al. Increased MRI activity and immune activation in two multiple sclerosis patients with the monoclonal anti-tumor necrosis factor antibody cA2. Neurology 1996; 47:1531-4. [PMID 8960740]
- Zheng l, Fisher G, Miller RE, Peschon J, Lynch DH, Lenardo MJ. Induction of apoptosis in mature T cells by tumor necrosis factor. Nature 1995; 377:348- 51. [PMID 7566090]
- Speiser DE, Sebzda E, Ohteki T, Bachmann MF, Pfeffer K, Mak TW, Ohashi PS. Tumor necrosis factor p55 mediates deletion of peripheral cytotoxic T lymphocytes in vivo. Eur J Immunol 1996; 26:3055-60. [PMID 8977304]
- Sytwu HK, Liblau RS, McDevitt HO. The role of Fas/Apo-1 (CD95) and TNF in antigen-induced programmed cell death in T cell receptor transgenic mice. Immunity 1996 5:17-30. [PMID 8758891]
- Verma BK, Fogarasi M, Szabo G. Downregulation of tumor necrosis factor alpha activity by acute ethanol treatment in human peripheral blood monocytes. J ClinImmunol 1993; 13:8. [PMID 8445046]
- Walsh K, Alexander G. Alcoholic liver disease. Postgrad Med J 2000; 76:280-6. [PMID 10775280]
- McClain CJ, Cohen DA. Increased tumor necrosis factor production by monocytes in alcoholic hepatitis. Hepatology 1989; 9:349-51. [PMID 2920991]
- Kamimura S, Tsukamoto H. Cytokine gene expression by Kuppfer cells in experimental alcoholic liver disease. Hepatology 1995; 21:1304-9. [PMID 7557885]
- Felver ME, Mezey E, McGuire M, Mitchell MC, Herlong HF, Veech GA, Veech RL. Plasma tumor necrosis factor alpha predicts long term survival in severe alcoholic hepatitis. Alcohol ClinExp Res1990; 14:255-9. [PMID 2190492]
- Bird GL, Sheron N, Goka AK, Alexander GJ, Williams RS. Increased tumor necrosis factor in severe alcoholic hepatitis. Ann Intern Med 1990; 112:917-20. [PMID 2339855]
- McClain C, Hill D, Schmidt J, Diehl AM. Cytokines and alcoholic liver disease. SeminLiv Dis 1993; 13:170-82. [PMID 8337603]
- Schilling PJ, Murray JL, Markowitz AB. Novel tumor necrosis factor toxic effects. Pulmonary hemorrhage and severe hepatic dysfunction. Cancer 1992; 69:256-60. [PMID 1309306]

