Keywords
alpha-Amylase; Cholecystokinin; Dietary Fats; Lipase; Time
Abbreviations
CCK: cholecystokinin; HFLCD: high-fat, low-carbohydrate diet
INTRODUCTION
It has been well-documented that the chronic intake of a high-fat diet is associated with hyperlipidemia, whose pathologic consequences are acute pancreatitis, atherosclerosis, obesity, non insulindependent diabetes mellitus and other disorders [1, 2, 3]. In addition, a high-fat diet may also be a significant risk factor in pancreatic cancer [4]. The elevation of blood lipid levels due to the ingestion of excess dietary fat can lead to the development of several disorders. The majority of fat digestion occurs in the upper intestine by lipase secreted from the exocrine pancreas.
It has been reported that the contents of pancreatic amylase, lipase and protease change in proportion to the dietary contents of their respective substrates, namely carbohydrates, fats and proteins [5, 6]. In addition, the synthesis, mRNA content and transcription of pancreatic lipase are regulated by dietary fat [7, 8]. The secretion of lipase from the exocrine pancreas is also dependent on the fat composition of the diet [9, 10]. However, considerable controversy exists concerning dietary fat and a consequent adaptation of the pancreatic enzymes. It has been demonstrated that dietary fat increases the lipase activity of the exocrine pancreas in mice, rats and pigs [7, 8, 9, 10]. Nevertheless, in other studies of conscious dogs, dietary fat failed to change the expression and secretion of pancreatic lipase [11, 12]. Although it has been well-documented that short-term fat given orally or intraduodenally induces the release of cholecystokinin (CCK) which is one of the potent mediators in pancreatic adaptation to dietary fat [13, 14, 15, 16], little is known about the adaptation of the exocrine pancreas to the chronic intake of dietary fat. Since increasing the fat component in diets results in the decrease of the carbohydrate component [5], pancreatic amylase could also be influenced by a reduced carbohydrate component.
Thus, the present study was undertaken to investigate the effects of very long-term highfat, low-carbohydrate feeding on the secretion of pancreatic amylase and lipase. The intracellular activity and the secretory activity of the pancreatic enzymes in the basal or in the CCK-stimulated states were compared in normal rats and high-fat, low-carbohydrate diet (HF-LCD) fed rats. Isolated pancreatic lobules of rats were utilized to test the enzyme secretion in this study in order to eliminate the possible influence of the extrinsic nerves and hormones on exocrine secretion.
MATERIALS AND METHODS
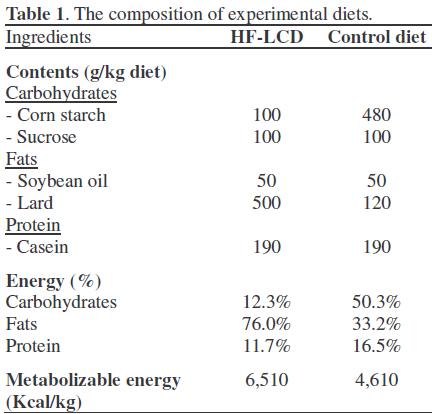
Animals and Diets
Prior to sacrifice, twenty-six male Sprague- Dawley rats (4 weeks of age, weighing 100- 150 g) were maintained for a period of 52 weeks in an environmentally-controlled room with a temperature of 23±2°C, relative humidity of 55±5% and a 12-h light:dark cycle with free access to water. The rats were randomly divided into two groups of 13 rats each. As shown in Table 1, the HF-LCD group was fed a diet containing 12.3% Kcal of carbohydrates, 76.0% Kcal of fat, and 11.7% Kcal of protein over a 48-week period. In the control group, the rats were fed a standard diet (containing 50.3% Kcal of carbohydrates, 33.2% Kcal of fat, and 16.5% Kcal of protein) for the same 48 weeks. The standard diets were prepared using a commercial source of AIN-93G purified rodent diet (Dyets, Bethelehem, PA, USA). The diets were mixed with cellulose (10 g/kg), L-cystine (3 g/kg), choline bitartrate (2.5 g/kg), AIG-93G vitamin mixture (10 g/kg), AIG-93G mineral mixture (35 g/kg), and tbutylhydroquinone (0.014 g/kg).
Preparation of the Pancreatic Lobules
After an overnight fast with free access to water, the animals were killed by cervical dislocation and the pancreatic lobules were obtained. The pancreatic lobules were prepared according to a method previously described [17]. In brief, the pancreas was quickly removed and immersed in modified calcium-free HEPES (25 mmol/L)-buffered Ringer solution with the following composition (Sigma-Aldrich, St. Louis, MO, USA): NaCl 113 mmol/L, HEPES 25 mmol/L, KCl 4.7 mmol/L, MgCl2 1.2 mmol/L, Dglucose 5.6 mmol/L, sodium pyruvate 5 mmol/L, sodium fumarate 5 mmol/L, sodium glutamate 5 mmol/L, and L-glutamine 2 mmol/L. The medium also contained 2% (v/v) essential amino acids, 1% (w/v) BSA, and 0.1% (w/v) soybean trypsin inhibitor. The solution was maintained at pH 7.4, while being continuously gassed with 100% O2 at 37°C. Whole pancreatic lobules were spread out by injecting calcium-free HEPES-Ringer solution gassed with 100% O2 into the loose connective tissue. The separated lobules, about 6 mg wet weight, were then obtained by excision. Four lobules were incubated in 4 mL of calcium-contained (2.5 mmol/L) HEPESbuffered Ringer solution gassed with 100% O2 after an equilibration period of 30 min.
Effect of Diets on the Intracellular Activities of Pancreatic Amylase and Lipase
The intracellular activities of alpha-amylase and lipase were measured in pancreatic lobules isolated from the control rats and the HF-LCD fed rats. The lobules were mechanically homogenized in 1% Triton X- 100 buffer (pH 8.0) containing (mmol/L) CaCl2 2.5 mmol/L, HEPES 25 mmol/L, and KCl 0.1 mmol/L. Alpha-amylase activity in the homogenate was determined according to method previously described, using starch as a substrate [18, 19]. Lipase activity was measured titrimetrically using lipase assay reagent (Sigma-Aldrich, St. Louis, MO, USA). Protein content in pancreatic tissue was measured using BCA protein assay reagent (Sigma-Aldrich, St. Louis, MO, USA).
Effects of Diets on the Secretory Activities of Pancreatic Amylase and Lipase in the Basal and in the CCK-stimulated States
To determine the secretory enzyme activity, 2 mL of the incubation medium was continuously collected every 30 min over the entire 90-min of the experimental period in an ice-chilled test tube and was immediately replaced with the same volume of fresh medium. The secretory activities of amylase and lipase in the basal state were measured in the pancreatic lobules isolated from the control rats and the HF-LCD fed rats without any secretagogues. To stimulate pancreatic enzyme secretion, synthetic sulfated CCK-8 (Sigma-Aldrich, St. Louis, MO, USA) at a concentration of 10 pmol/L was introduced into the culture medium, and the concentration was then maintained by replacement with 2 mL of fresh medium containing 10 pmol/L of CCK after the sample collection every 30 min. To calculate the enzyme activity initially present within the lobules, alpha-amylase and lipase activity not only in the incubation medium but also in the tissue were determined.
ETHICS
All the animals involved in this experiment received humane care according to the criteria outlined in the “Guide for the Care and Use of Laboratory Animal (1996)” prepared by the National Academy of Sciences.
STATISTICS
All results are presented as means±SE. The intracellular activities of amylase and lipase were defined as amylase and lipase activity per 1 mg of protein in the pancreatic homogenate. The secretory activities of amylase and lipase were defined as U/90 min/mg protein, which is the enzyme activity cumulatively secreted over the entire 90-min period in the medium per 1 mg of protein initially present in the lobules, and as a % total, which is enzyme activity cumulatively secreted over the entire 90-min period in the medium per enzyme activity initially present within lobules. The protein amount and the enzyme activity initially present in the lobules were calculated from the sum of the secreted value and the retained value. The data were analyzed using the Student's t test by means of the SPSS version 9.0 for Windows. Differences were considered significant when the two-tailed P value was less than 0.05.
RESULTS
Intracellular Activities of Pancreatic Amylase and Lipase
The mean body weights of the rats prior to being fed the control diet and the HF-LCD were 120±6 g and 121±7 g, respectively. After 48 weeks, their body weights increased to 408±22 g and 499±28 g in the control group and in the HF-LCD group, respectively. Significantly greater weight gain (P=0.016) was observed in the HF-LCD group. As shown in Figure 1a, the intracellular alphaamylase activity in the pancreatic lobules isolated from HF-LCD fed rats was 71.6±5.0 U/mg protein, which was significantly lower (P=0.007) than that in control rats (95.0±5.8 U/mg protein). However, the intracellular lipase activity (Figure 1b) in the HF-LCD fed rats (329±21 U/mg protein) was not significantly different (P=0.362).
Figure 1. The intracellular activities of amylase (a.)
and lipase (b.) in pancreatic lobules isolated from
control rats and high-fat, low-carbohydrate diet (HFLCD)
fed rats. Each bar represents the mean±SE of
data obtained from 13 experiments. The intracellular
amylase activity in HF-LCD fed rats was significantly
lower than that in control rats. The intracellular lipase
activity was not significantly changed by HF-LCD
feeding.
Secretory Pancreatic Enzyme Activities in the Basal and in the CCK-stimulated States
The pancreatic lobules of the control rats spontaneously secreted amylase and lipase in the basal state at a rate of 11.1±1.5 U/90 min/mg protein and 25.4±2.7 U/90 min/mg protein, respectively. As shown in Figure 2, the lobules of the HF-LCD fed rats spontaneously secreted amylase at a rate of 6.85±1.47 U/90 min/mg protein, which was notably lower (P=0.030) than that in control rats while they secreted lipase at a rate of 27.7±2.9 U/90 min/mg protein, which was similar to that in control rats (P=0.342). However, when secretory enzyme activity was illustrated as the percentage released, the secretory activities of amylase and lipase in basal state of HF-LCD group were not significantly different from the control group (Table 2).
Figure 2. The secretory activities of amylase (a.) and
lipase (b.) in the basal and in the CCK-stimulated
states (10 pmol/L) over the entire 90-min period in
pancreatic lobules isolated from control rats and HFLCD
fed rats. Each bar represents the mean±SE of data
obtained from 13 experiments. The secretory amylase
activities in thebasal and in the CCK-stimulated states
in HF-LCD fed rats were significantly lower than those
in control rats. The secretory lipase activities in the
basal and in the CCK-stimulated states were not
significantly changed by HF-LCD feeding.
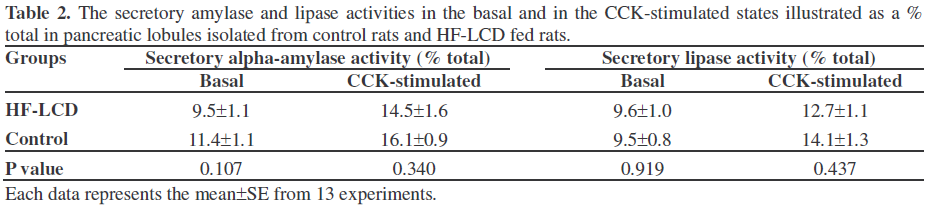
As shown in Figure 3, the CCK-stimulated pancreatic amylase secretion was dosedependent. Significance (P<0.001) was observed beginning with 10 pmol/L of CCK, and maximal effect was observed in 1 nmol/L of CCK. CCK at a concentration of 10 pmol/L significantly increased the secretory activities of amylase and lipase in HF-LCD fed rats to 10.5±1.6 U/90 min/mg protein and 36.2±3.0 U/90 min/mg protein, respectively (Figure 2). In control rats, 10 pmol/L of CCK also increased the secretory activities of amylase and lipase to 16.1±1.7 U/90 min/mg protein and 35.3±2.7 U/90 min/mg protein, respectively. However, the secretory amylase activity in the CCK-stimulated state of HFLCD fed rats was lower (P=0.022) than that of control rats, despite the fact that the secretory lipase activity was not different in the two groups (P=0.811). Moreover, when the secretory enzyme activity was illustrated as the percentage released, the secretory activities of amylase and lipase in the CCKstimulated state of the HF-LCD group were not significantly different from the control group (Table 2).
Figure 3. Dose-response curve of the effect of
cholecyctokinin (CCK) on secretory amylase activity
in pancreatic lobules isolated from control rats. a. Time
courses of the secretory amylase activity stimulated by
CCK. b. Cumulative amylase activity secretory over
the entire 90-min period. Each value represents the
mean±SE of data obtained from 6 experiments. CCK
significantly enhanced the secretory amylase activity
from the concentration of 10 pmol/L, and maximal
effect was observed at 1 nmol/L of CCK.
DISCUSSION
In the present study, we first report that the intracellular and the secretory activity of amylase were reduced by very long-term (48 weeks) high-fat, low-carbohydrate feeding. In our study, we used a control diet containing 50.3% Kcal of carbohydrates and a HF-LCD containing 12.3% Kcal of carbohydrates. In general, increasing the fat component in the diets results in a decrease in the carbohydrate component [5]. It has been well-established that high-fat, low-carbohydrate feeding results in a decrease of the mRNA expression of pancreatic amylase [20, 21] and highcarbohydrate feeding results in an increase of the mRNA expression of pancreatic amylase in rats [22]. These results strongly indicate that high-fat feeding may reduce pancreatic amylase synthesis due to a low-carbohydrate component present in the duodenum for an extended period of time, namely a long-term adaptation.
In in vitro studies, a high-fat diet decreased the basal or the CCK-stimulated secretion of amylase, and a high-carbohydrate diet increased them in rats [10, 21, 23, 24]. In our study, the secretory amylase activities in the basal and in the CCK-stimulated states were lower in HF-LCD fed rats than in control diet fed rats. However, when the secretory amylase activity was illustrated as the percentage released, the secretory amylase activities in the basal and in the CCKstimulated states of high-fat feeding rats were not significantly different from those of control diet fed rats. Therefore, the reduced amylase activities in the basal and in the CCK-stimulated states, illustrated as U/90 min/mg protein, may be due to the reduced intracellular amylase activity rather than to altered sensitivity of the pancreatic lobules to CCK. These results suggest that the synthesis of pancreatic amylase may adapt to the reduced load of carbohydrates for digestion in the small intestine.
Generally, increasing dietary fat stimulates the synthesis [7, 25], mRNA content [8, 26] and transcription [27] of pancreatic lipase. However, this adaptation of lipase seems to be time-dependent since controversial results have been observed in relation to the length of time of dietary fat feeding. Some studies in dogs showed that dietary fat feeding for only 8 days failed to change the expression of pancreatic lipase [11, 12]. However, other studies showed that lipase was sensitive to high-fat feeding for 4 weeks [8, 10]. It has recently been reported that elevated lipase mRNA expression is sustained for 8 weeks but returned to normal levels by 16 weeks in high-fat feeding mice [7]. Similarly, intracellular lipase activity was also unchanged by long-term HF-LCD feeding despite a reduction in the intracellular amylase activity in the present study. Although the mechanism of unchanged intracellular lipase activity is not known, it seems that an adaptation of intracellular lipase activity to a HF-LCD diet may need appropriate feeding duration, and may not be sustained for a long time.
It is known that dietary fat enhances the pancreatic secretion of lipase [9, 10, 16]. CCK is thought to be one of the potent mediators of an adaptation of pancreatic lipase, since short-term fat given orally or intraduodenally has been known to induce the release of CCK which is one of the potent stimulators of enzyme secretion of the pancreas [13, 14, 27, 28, 29]. Interestingly, it was reported that there were no differences in plasma CCK secretion between lean and obese people in a fasting state as well as in response to the ingestion of a mixed meal [30, 31]. Moreover, CCK-deficient mice clearly demonstrated that CCK was not required for the long-term protein-induced adaptation of pancreatic protease [32], despite the fact that CCK has been considered the primary mediator of dietary protein-induced changes in the pancreas [5]. Although there were conflicting reports regarding the role of CCK on the adaptation of the production of pancreatic enzymes, we failed to modify the secretory lipase activities in the basal and in the CCK-stimulated states by long-term HFLCD feeding as compared to control diet feeding. We used CCK at a concentration of 10 pmol/L which is a relatively lower, but physiological [27, 28, 29], concentration as compared to other studies; thus, the secretory lipase activity may be unchanged in this study. This result may also be an age-related phenomenon because there is a reduction in lipase secretion in older humans [33, 34]. Therefore, further study about pancreatic lipase secretion with a higher dose of CCK or other secretagogues is needed to elucidate the adaptive response of pancreatic lipase to HFLCD feeding.
From the above results, we conclude that reduction of intracellular and secretory amylase activity may be a long-term adaptive response of the pancreas to a reduced carbohydrate component, and this long-term adaptation of the pancreatic secretion of amylase may be due to the changes in the intracellular amylase activity rather than to the altered sensitivity of the acinar to CCK.
References
- Sumiyoshi M, Sakanaka M, Kimura Y. Chronic intake of high-fat and high-sucrose diets differentially affects glucose intolerance in mice. J Nutr 2006; 136:582-7. [PMID 16484528]
- Hennig B, Toborek M, McClain CJ. High-energy diets, fatty acids and endothelial cell function: implications for atherosclerosis. J Am Coll Nutr 2001; 20:97-105. [PMID 11349944]
- Bray GA, Gray DS. Obesity. Part I-Pathogenesis. West J Med 1988; 149:429-41. [PMID 3067447]
- Zhang J, Go VL. High fat diet, lipid peroxidation, and pancreatic carcinogenesis. Adv Exp Med Biol 1996; 399:165-72. [PMID 8937556]
- Brannon PM. Adaptation of the exocrine pancreas to diet. Annu Rev Nutr 1990; 10:85-105. [PMID 2200477]
- Kern HF, Rausch U, Scheele GA. Regulation of gene expression in pancreatic adaptation to nutritional substrates or hormones. Gut 1987; 28:89-94. [PMID 3319815]
- Rippe C, Berger K, Mei J, Lowe ME, Erlanson- Albertsson C. Effect of long-term high-fat feeding on the expression of pancreatic lipases and adipose tissue uncoupling proteins in mice. Pancreas 2003; 26:36-42. [PMID 12604926]
- Ricketts J, Brannon PM. Amount and type of dietary fat regulate pancreatic lipase gene expression in rats. J Nutr 1994; 124:1166-71. [PMID 8064366]
- Hedemann MS, Pedersen AR, Engberg RM. Exocrine pancreatic secretion is stimulated in piglets fed fish oil compared with those fed coconut oil or lard. J Nutr 2001; 131:3222-6. [PMID 11739870]
- Chowdhury P, Nishikawa M, Blevins GW Jr, Rayford PL. Response of rat exocrine pancreas to highfat and high-carbohydrate diets. Proc Soc Exp Biol Med 2000; 223:310-5. [PMID 10719845]
- Manas M, Yago MD, Quiles JL, Huertas JR, Martinez-Victoria E. Absence of rapid adaptation of the exocrine pancreas of conscious dogs to diets enriched in fat or carbohydrates. Arch Physiol Biochem 1996; 104:819-25. [PMID 9127677]
- Yago MD, Martinez-Victoria E, Huertas JR, Manas M. Effects of the amount and type of dietary fat on exocrine pancreatic secretion in dogs after different periods of adaptation. Arch Physiol Biochem 1997; 105:78-85. [PMID 9224549]
- Spannagel AW, Nakano I, Tawil T, Chey WY, Liddle RA, Green GM. Adaptation to fat markedly increases pancreatic secretory response to intraduodenal fat in rats. Am J Physiol 1996; 270:G128-35. [PMID 8772510]
- Hildebrand P, Petrig C, Burckhardt B, Ketterer S, Lengsfeld H, Fleury A, et al. Hydrolysis of dietary fat by pancreatic lipase stimulates cholecystokinin release. Gastroenterology 1998; 114:123-9. [PMID 9428226]
- Marion J, Rome V, Savary G, Thomas F, Le Dividich J, Le Huerou-Luron I. Weaning and feed intake alter pancreatic enzyme activities and corresponding mRNA levels in 7-d-old piglets. J Nutr 2003; 133:362-8. [PMID 12566468]
- Jakob S, Mosenthin R, Zabielski R, Rippe C, Winzell MS, Gacsalyi U, et al. Fats infused intraduodenally affect the postprandial secretion of the exocrine pancreas and the plasma concentration of cholecystokinin but not of peptide YY in growing pigs. J Nutr 2000; 130:2450-5. [PMID 11015471]
- Scheele GA, Palade GE. Studies on the guinea pig pancreas. Parallel discharge of exocrine enzyme activities. J Biol Chem 1975; 250:2660-70. [PMID 1123325]
- Park HS, Park HJ. Effects of gamma-aminobutyric acid on secretagogue-induced exocrine secretion of isolated, perfused rat pancreas. Am J Physiol Gastrointest Liver Physiol 2000; 279:G677-82. [PMID 11005753]
- Rick W, Stegbauer HP. Alpha-Amylase: Measurement of reducing groups. In: Bergermeyer HU, ed. Methods of Enzymatic Analysis. Volume 2. 2nd ed, Weinheim; Verlag Chemie, 1974:885-915.
- Tsai A, Cowan MR, Johnson DG, Brannon PM. Regulation of pancreatic amylase and lipase gene expression by diet and insulin in diabetic rats. Am J Physiol 1994; 267:G575-83. [PMID 7524347]
- Takaori K, Blevins GT Jr, Nishikawa M, Chowdhury P, Rayford PL. Effects of diet on cholecystokinin-stimulated amylase secretion by pancreatic Acini and amylase mRNA levels in rat pancreas. Pancreas 1995; 11:402-7. [PMID 8532658]
- Giorgi D, Bernard JP, Lapointe R, Dagorn JC. Regulation of amylase messenger RNA concentration in rat pancreas by food content. EMBO J 1984; 3:1521- 4. [PMID 6204864]
- Sommer H, Kasper H. Dietary effects on pancreatic exocrine function. Experiments on the isolated perfused rat pancreas. Ann Nutr Metab 1981; 25:381-8. [PMID 6174069]
- Holler B, Ogami Y, Zabel-Langhennig A, Tillack D, Engeland K, Keim V, Mossner J. Role of prostaglandins in regulation of pancreatic enzyme secretion by various diets. Dig Dis Sci 2001; 46:289- 95. [PMID 11281177]
- Birk RZ, Brannon PM. Regulation of pancreatic lipase by dietary medium chain triglycerides in the weanling rat. Pediatr Res 2004; 55:921-6. [PMID 15071157]
- Wicker C, Puigserver A. Expression of rat pancreatic lipase gene is modulated by a lipid-rich diet at a transcriptional level. Biochem Biophys Res Commun 1990; 166:358-64. [PMID 1689152]
- Watanabe S, Lee KY, Chang TM, Berger-Ornstein L, Chey WY. Role of pancreatic enzymes on release of cholecystokinin-pancreozymin in response to fat. Am J Physiol 1988; 254:G837-42. [PMID 3377083]
- Green GM, Taguchi S, Friestman J, Chey WY, Liddle RA. Plasma secretin, CCK, and pancreatic secretion in response to dietary fat in the rat. Am J Physiol 1989; 256:G1016-21. [PMID 2735407]
- Park HS, Park IS, Lee YL, Kwon HY, Park HJ. Effects of intrapancreatic neuronal activation on cholecystokinin-induced exocrine secretion of isolated perfused rat pancreas. Pflugers Arch 1999; 437:511-6. [PMID 10089562]
- Lieverse RJ, Masclee AA, Jansen JB, Lamers CB. Plasma cholecystokinin and pancreatic polypeptide secretion in response to bombesin, meal ingestion and modified sham feeding in lean and obese persons. Int J Obes Relat Metab Disord 1994; 18:123-7. [PMID 8148926]
- Folsch UR, Dreessen UW, Talaulicar M, Willms B, Creutzfeldt W. Effect of long-term fasting of obese patients on pancreatic exocrine function, gastrointestinal hormones and bicarbonate concentration in plasma. Z Gastroenterol 1984; 22:357- 64. [PMID 6207670]
- Lacourse KA, Swanberg LJ, Gillespie PJ, Rehfeld JF, Saunders TL, Samuelson LC. Pancreatic function in CCK-deficient mice: adaptation to dietary protein does not require CCK. Am J Physiol 1999; 276:G1302-9. [PMID 10330022]
- Vellas B, Balas D, Moreau J, Bouisson M, Senegas-Balas F, Guidet M, Ribet A. Exocrine pancreatic secretion in the elderly. Int J Pancreatol 1988; 3:497-502. [PMID 2464656]
- Laugier R, Bernard JP, Berthezene P, Dupuy P. Changes in pancreatic exocrine secretion with age: pancreatic exocrine secretion does decrease in the elderly. Digestion 1991; 50:202-11. [PMID 1812045]




