Keywords
Cadherins; Pancreas; Pancreatic Neoplasms
Abbreviations
SPT: solid pseudopapillary tumor; TMA: tissue microarray analysis
INTRODUCTION
Decrease in or loss of E-cadherin is considered a molecular event signaling dysfunction of the cell-cell adhesion system, thereby triggering invasion and metastasis in many malignancies. In addition, alterations in the cadherin-catenin system may also be involved in tumorigenesis. E-cadherin and beta-catenin, components of the Wnt signal transduction pathway, may serve as a common switch in central processes that regulate cellular differentiation and growth. Abnormalities of expression of beta-catenin protein and in the gene have been reported in solid pseudopapillary tumors (SPT) of the pancreas, and are considered a recurring, consistent finding associated with these lesions [1, 2]. SPT are rare neoplasms with a low malignant potential occurring most frequently in young women. Histologically, it is a solid and cystic tumor with a prominent vascular network and degenerative pseudopapillae [3]. Often times they may resemble endocrine tumors of the pancreas very closely because of the vascular network and relatively uniform cytological appearance. These tumors are composed of small uniform cells with nuclei that can show grooves and irregular contours.
The neoplastic cells in SPT are consistently vimentin, CD10 and CD56-positive and nuclear beta-catenin expression is also a wellrecognized feature. Some cases express focal positivity for alpha-1-antitrypsin, alpha-1- antichymotrypsin, neuron-specific enolase and synaptophysin.
In view of the association between E-cadherin and beta-catenin, the purpose of this study was to examine expression of E-cadherin in a series of SPT using a tissue microarray and to correlate immunoexpression with clinicopathological parameters.
MATERIALS AND METHODS
Eighteen cases of SPT of the pancreas were utilized for tissue microarray analysis (TMA): 10 were retrieved from the archives of the Department of Pathology, University Heath Network, Toronto, and 8 from the Department of Pathology, McMaster University, Hamilton.
Tissue Microarray Analysis Construction
For each case, 4 to 14 cores measuring 1.0 mm each were drilled from the blocks. Tissue cores from normal pancreas were used as controls and for orientation purposes.
Size of tumor, necrosis, gross invasiveness/ demarcation, lymphovascular invasion, lymph node involvement and liver metastasis were recorded. The mitotic count, expressed per 10 high powered fields (HPFs) was assessed.
Immunohistochemistry
All the TMA blocks were stained with a panel of antibodies used in the routine work-up of SPTs, in addition to including beta-catenin and E-cadherin which form the main thrust of this study (Table 1). Tissue sections were cut using sterile disposable microtome blades on a rotary microtome. They were floated on a water bath (50°C) and picked up on poly-Llysine coated glass slides. The sections were baked on a hot plate at 60°C for 20 minutes. Deparaffinization was carried out in 2 changes of xylene for 5 minutes each. This was followed by rehydration in 2 changes of absolute ethanol for 3 minutes each and 2 changes of 95% ethanol for 3 minutes each before a stay of 5 minutes under running tap water. For the antibodies requiring microwave heat retrieval, the sections were processed to unmask antigens by conventional microwave heating in 0.01 M sodium citrate retrieval buffer (Table 1). The other heat retrieval method is indicated in Table 1.

Appropriate positive controls (cases known to be positive for all antibodies used in the study) were run in parallel. SPTs were regarded as positive for a particular antibody if the tumor cells displayed a similar staining pattern and intensity as the known positive control. Specifically, membranous immunoreactivity for E-cadherin/beta-catenin was considered normal; any reduction or loss of this pattern of immunoreactivity compared to normal tissue, together with nuclear and/or cytoplasmic staining was considered as abnormal immunoexpression.
ETHICS
Written informed consent was obtained from each patient and the study protocol conforms to the ethical guidelines of the "World Medical Association Declaration of Helsinki - Ethical Principles for Medical Research Involving Human Subjects" adopted by the 18th WMA General Assembly, Helsinki, Finland, June 1964, as revised in Tokyo 2004, as reflected in a priori approval by the appropriate institutional review committee.
STATISTICS
Means, ranges, and frequencies were evaluated as descriptive statistics. The Mann- Whitney U-test was applied to compare cases with and without metastases. Two-tailed P values less than 0.05 were considered significant. Data were analyzed by using the SPSS for Windows (version 13.0) package.
RESULTS
Patient and tumor characteristics of the 18 cases of solid pseudopapillary tumor of the pancreas are shown in Table 2. Sixteen of the 18 cases were in female patients, 2 were males, ranging in age from 11 to 62 years (average: 38.0 years). Fifteen tumors arose in the tail/body of the pancreas, while 3 (16.7%) were located in the head. The size of SPT ranged from 2 to 17 cm, with a mean size of 6.3 cm. Four cases (22.2%) had synchronous liver metastases; in addition one patient had a liver recurrence one year later after the resection of the primary tumor. All the tumors were macroscopically well circumscribed, but unencapsulated. On sectioning, they displayed solid areas with focally hemorrhagic and unior multilocular cystic spaces, filled with fluid ranging from straw- to red-colored. Histologically, all SPT studied showed a proliferation of uniform neoplastic cells arranged in sheets, nests and cords and characterized by eosinophilic and cytoplasmic vacuoles that sometimes indented grooved and vesicular nuclei (Figure 1). Nucleoli were inconspicuous. Periodic acid Schiff (PAS) and PAS following diastase digestion (PASD) positive hyaline globules were variably found in the cytoplasm and/or in the intercellular spaces. The mitotic activity was minimal and abnormal figures were not discerned. The average mitotic count was 1 mitotic figure per 10 HPF (range: 0-3) across all tumors. Degenerative changes were variably present and pseudo-papillae consisted of a fibrovascular core lined by poorly cohesive neoplastic cells (Figure 2). Cholesterol clefts, foamy macrophages and foreign body giant cells were seen. In areas of hemorrhage there was degeneration and deposition of hemosiderin. Although tumors are well demarcated from the surrounded pancreas, a true capsule was lacking and small projections of the tumor extended into the normal pancreatic parenchyma in a smooth, non-infiltrative manner. Lymphovascular or lymph node involvement was not seen in any case.

Figure 1. High power illustration of the cytology of a
solid pseudopapillary tumor with clear cells, many of
which contain indented and irregularly shaped nuclei
(H&E x400).
Figure 2. The typical appearance of pseudo-papillary
structures where the cells have lost cohesion (H&E
x200).
Immunohistochemistry
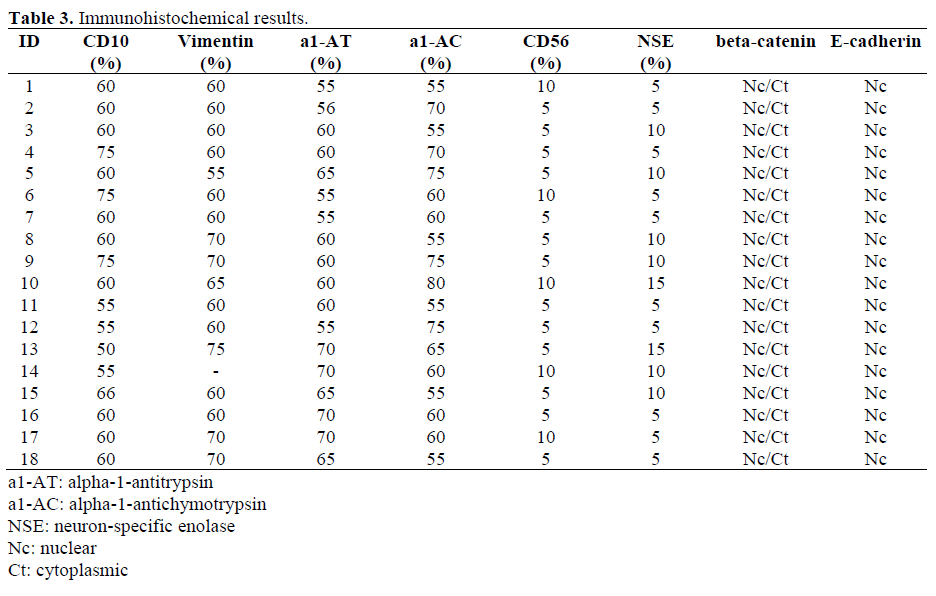
Immunohistochemical results are shown in Table 3. All 18 cases contained 50% or more of tumor cells that were CD10 (cytoplasmic and often dot-like) (mean: 61.4%, range 50- 75%), vimentin (mean: 63.2%, range 55-75%; 17 cases were available), alpha-1-antitrypsin (mean: 61.7%, range 55-70%) and alpha-1- antichymotrypsin (mean: 63.3%, range 55- 80%) positive. At least focal positivity was seen in all cases for CD56 and the range of immunopositivity was from 5 to 10% of tumor cells labeling (mean 7.8%). NSE staining was cytoplasmic (mean: 7.8%, range 5-15%). All cases were negative for chromogranin and synaptophysin.
All 18 cases displayed complete loss of membrane expression for beta-catenin; only cytoplasmic and nuclear localization of the protein was present in all tumor cells (Figure 3).
Figure 3. Intense nuclear labeling for beta-catenin
(anti-beta-catenin x400).
Similarly, E-cadherin protein was localized to the nucleus in all 18 cases, with loss of the characteristic membranous decoration of cells (Figure 4).
Figure 4. The tumour cells in solid pseudopapillary
tumor show loss of membrane staining for E-cadherin;
instead E-cadherin is now located in the nucleus (anti-
E-cadherin x400).
There was no significant differences in the immunohistochemical results between the 4 cases with metastases and those that were confined to the pancreas (CD10, P=0.128: mean 57.5%, range 55-60% vs. mean 62.6%, range 50-75%; vimentin, P=0.776: mean 61.7%, range 60-65%, n=3 vs. mean 63.6%, range 55-75%; alpha-1-antitrypsin, P=0.827: mean 61.3%, range 55-70% vs. mean 61.9%, range 55-70%; alpha-1-antichymotrypsin, P=0.661: mean 62.5%, range 55-80% vs. mean 63.6%, range 55-75%; neuron-specific enolase, P=0.633: mean 8.8%, range 5-15% vs. mean 7.5%, range 5-15%) a part from CD56 that was significantly (P=0.020) higher in metastases cases (mean 8.8%, range 5-10% vs. mean 5.7%, range 5-10%). The significance of CD56 staining lesions that metastasize is conjectural but may serve as a marker for lesions with metastatic potential. However, more cases need to be examined to validate this.
DISCUSSION
SPT is a rare pancreatic tumor, constituting 1- 2% of exocrine pancreatic tumors and about 5% of cystic pancreatic tumors, and its origin is still unresolved and controversial although a ductal origin has been favoured [3]. Although, it is a low-grade malignant tumor with a good prognosis after surgical excision, widespread liver metastases and vascular invasion have been reported [4], but invasion into surrounding organs such as the spleen and duodenum is rare [3].
Histologically, it is quite characteristic if not diagnostic, in most cases, however, there are variants such as the predominantly solid cases and even a clear cell variant that may cause mimicry of other tumours [5].
SPT is consistently immunoreactive for CD10, vimentin and CD56 and, frequently so for alpha-1-antitrypsin and alpha-1- antichymotrypsin. Sometimes immunoreactivity for synaptophysin is seen.
E-cadherin and beta-Catenin, two key components of the Wnt signal transduction pathway, are involved in cellular differentiation and growth [6]. The Ecadherin gene (CDH 1) is located at locus 16q22.1 and produces a 120 kDa protein. Ecadherin localizes to zonula adherens junctions that are adherens junctions typically seen in epithelial cells. Normal E-cadherin expression plays a key role in the maintenance of epithelial integrity and polarized function [7, 8]. Reduced E-cadherin expression was shown to promote epithelial cell invasiveness, dedifferentiation and metastases in various human carcinomas, supporting a role for this protein as an “invasion suppressor molecule” [9, 10, 11].
E-cadherin is a calcium-dependent transmembrane glycoprotein, involved in the cellcell interaction and is localized to the basolateral surface of the epithelial cells. It has a cytoplasmic domain, a single-pass transmembrane domain and an extracellular domain that consists of five tandemlyrepeated cadherin-motifs sub-domains with putative calcium binding sites. The extracellular N-terminal end is essential for the homophilic calcium-dependent cell-cell adhesion [12]. Although cadherin binding is not a direct activator of Wnt-Wingless signal transduction, recently it has been demonstrated that classical cadherins do function as receptors in GTPase signaling [13, 14]. In particular, the small GTPases of the Rho family have been identified as part of a membrane-local signaling process that regulates cell actin dynamics, cell motility and adherens junction assembly in response to cadherin adhesion [15, 16]. It is thought that cadherin adhesion may influence the precise sites at the plasma membrane where Rho family signaling occurs. Thus, the loss of cadherin-mediated cell-cell adhesion is an important event that has many far-reaching consequences for the acquisition of the invasive phenotype and tumour progression. The cytoplasmic domain of E-cadherin interacts with the catenin molecules that mediate its binding to the actin cytoskeleton. E-cadherin binds to either beta-catenin or gamma-catenin, but not to alpha-catenin [12]. Reduction in cell adhesion is considered a pivotal step in tumor metastasis and may be achieved by several mechanisms involving the E-cadherin-catenin complex, including the reduction or loss of E-cadherin expression, mutation or reduced transcription of the genes of the constituent molecules, redistribution of E-cadherin to different sites within the cell, shedding of E-cadherin, and competition for binding sites from other proteins [12].
beta-catenin is an integral component of the E-cadherin complexes at intercellular adherens junctions, and also recruits chromatin-remodeling complexes to activate transcription in the nucleus. The cytoplasmic tail of E-cadherin protein binds to betacatenin that in turn complexes with alphacatenin to form the E-cadherin-catenin unit. The presence of some tumours with normal expression of beta-catenin but abnormalities of E-cadherin may suggest that E-cadherin dysregulation may also be independent of beta-catenin. Alterations of E-cadherin are more frequently seen than those of beta catenin in tumours in general, but there still appears to be a correlation between the two proteins.
Recently, nuclear beta-catenin has been found to be present in all reported cases of SPT [2] and has been associated with activating mutation, (34)Gly(GGA) to Arg(AGA), in exon 3 of the beta-catenin gene [2, 17]. All the tumors in our series showed cytoplasmic and nuclear immunoreactivity for beta-catenin. In addition, we found, and report for the first time, that SPT also displayed consistent nuclear localization of E-cadherin protein associated with a concomitant loss of membrane staining of E-cadherin. Built-in positive control of normal ducts and pancreatic parenchyma still showed prominent membrane decoration with Ecadherin.
Although E-cadherin does not contain any identifiable nuclear localization signal sequence, the cleaved or short fragment of Ecadherin can translocate to the nucleus. The exact mechanism by which E-cadherin is over-expressed, stabilized or translocates to the nucleus is not clear. One possibility is that the cleaved E-cadherin, which normally is degraded by proteasome degradation, due to abnormality in the proteosomal pathway accumulates and subsequently translocates to the nucleus where it might participate in regulation of gene expression. Other studies suggest that E-cadherin modulates Wntdependent gene expression by regulating the availability of beta-catenin [18]. In malignant melanoma loss of E-cadherin leads to upregulation of NF-kappa beta activity [19]. E-cadherin dependent cell-cell adhesion has been shown to control the nuclear abundance of the nuclear transcription factors such as apolipoprotein A-IV [19]. Cell-matrix and cell-cell adhesion play a central role in the control of cell proliferation, differentiation, and gene expression and E-cadherin appears to play a key role in these processes in epithelial cells. Some studies suggest that loss of E-cadherin expression does not induce Wnt signaling, while in other studies a role of Ecadherin in modulation of Wnt-dependent gene expression has been suggested [19, 20, 21].
In view of the close association between betacatenin and E-cadherin, we speculate that the nuclear localization of E-cadherin is related to beta-catenin abnormalities in SPT. Given that beta-catenin gene abnormalities are common in SPT, it is possible that the E-cadherincatenin unit is perturbed by such mutations leading to loss of E-cadherin function. Although E-cadherin has a role in both tumour and invasion suppression, its nuclear localization in SPT is not correlated with metastasis.
References
- Abraham SC, Lee JH, Hruban RH, Argani P, Furth EE, Wu TT. Molecular and immunohistochemical analysis of intraductal papillary neoplasms of the biliary tract. Hum Pathol 2003; 34:902-10. [PMID 14562286]
- Min Kim S, Sun CD, Park KC, Kim HG, Lee WJ, Choi SH. Accumulation of beta-catenin protein, mutations in exon-3 of the beta-catenin gene and a loss of heterozygosity of 5q22 in solid pseudopapillary tumor of the pancreas. J Surg Oncol 2006; 94:418-25. [PMID 16967453]
- Santini D, Poli F, Lega S. Solid-papillary tumors of the pancreas: histopathology. JOP. J Pancreas (Online) 2006; 7:131-6. [PMID 16407635]
- Saiura A, Umekita N, Matsui Y, Maeshiro T, Miyamoto S, Kitamura M, Wakikawa A. Successful surgical resection of solid cystic tumor of the pancreas with multiple liver metastases and a tumor thrombus in the portal vein. Hepatogastroenterology 2000; 47:887- 9. [PMID 10919054]
- Albores-Saavedra J, Simpson KW, Bilello SJ. The clear cell variant of solid pseudopapillary tumor of the pancreas: a previously unrecognized pancreatic neoplasm. Am J Surg Pathol 2006; 30:1237-42. [PMID 17001153]
- Willert K, Jones KA. Wnt signaling: is the party in the nucleus? Genes Dev 2006; 20:1394-404. [PMID 16751178]
- Larue L, Ohsugi M, Hirchenhain J, Kemler R. Ecadherin null mutant embryos fail to form a trophectoderm epithelium. Proc Natl Acad Sci USA 1994; 91:8263-7. [PMID 8058792]
- Nelson WJ, Shore EM, Wang AZ, Hammerton RW. Identification of a membrane-cytoskeletal complex containing the cell adhesion molecule uvomorulin (E-cadherin), ankyrin, and fodrin in Madin-Darby canine kidney epithelial cells. J Cell Biol 1990; 110:349-57. [PMID 2153683]
- Dorudi S, Sheffield JP, Poulsom R, Northover JM, Hart IR. E-cadherin expression in colorectal cancer. An immunocytochemical and in situ hybridization study. Am J Pathol 1993; 142:981-6. [PMID 7682766]
- Gamallo C, Palacios J, Suarez A, Pizarro A, Navarro P, Quintanilla M, Cano A. Correlation of Ecadherin expression with differentiation grade and histological type in breast carcinoma. Am J Pathol 1993; 142:987-93. [PMID 7682767]
- Vleminckx K, Vakaet L Jr, Mareel M, Fiers W, van Roy F. Genetic manipulation of E-cadherin expression by epithelial tumor cells reveals an invasion suppressor role. Cell 1991; 66:107-19. [PMID 2070412]
- Beavon IR. The E-cadherin-catenin complex in tumour metastasis: structure, function and regulation. Eur J Cancer 2000; 36:1607-20. [PMID 10959047]
- Charrasse S, Meriane M, Comunale F, Blangy A, Gauthier-Rouviere C. N-cadherin-dependent cell-cell contact regulates Rho GTPases and beta-catenin localization in mouse C2C12 myoblasts. J Cell Biol 2002; 158:953-65. [PMID 12213839]
- Noren NK, Liu BP, Burridge K, Kreft B. p120 catenin regulates the actin cytoskeleton via Rho family GTPases. J Cell Biol 2000; 150:567-80. [PMID 10931868]
- Braga VM, Machesky LM, Hall A, Hotchin NA. The small GTPases Rho and Rac are required for the establishment of cadherin-dependent cell-cell contacts. J Cell Biol 1997; 137:1421-31. [PMID 9182672]
- Kaibuchi K, Kuroda S, Fukata M, Nakagawa M. Regulation of cadherin-mediated cell-cell adhesion by the Rho family GTPases. Curr Opin Cell Biol 1999; 11:591-6. [PMID 10508646]
- Nishimori I, Kohsaki T, Tochika N, Takeuchi T, Minakuchi T, Okabayashi T, et al. Non-cystic solidpseudopapillary tumor of the pancreas showing nuclear accumulation and activating gene mutation of betacatenin. Pathol Int 2006; 56:707-11. [PMID 17040296]
- Kuphal F, Behrens J. E-cadherin modulates Wntdependent transcription in colorectal cancer cells but does not alter Wnt-independent gene expression in fibroblasts. Exp Cell Res 2006; 312:457-67. [PMID 16368435]
- Kuphal S, Poser I, Jobin C, Hellerbrand C, Bosserhoff AK. Loss of E-cadherin leads to upregulation of NFkappaB activity in malignant melanoma. Oncogene 2004; 23:8509-19. [PMID 15378016]
- Peignon G, Thenet S, Schreider C, Fouquet S, Ribeiro A, Dussaulx E, et al. E-cadherin-dependent transcriptional control of apolipoprotein A-IV gene expression in intestinal epithelial cells: a role for the hepatic nuclear factor 4. J Biol Chem 2006; 281:3560- 8. [PMID 16338932]
- Herzig M, Savarese F, Novatchkova M, Semb H, Christofori G. Tumor progression induced by the loss of E-cadherin independent of beta-catenin/Tcfmediated Wnt signaling. Oncogene 2006. Epub ahead of print. [PMID 17043652]





