Keywords
Autonomic Pathways; Enteric Nervous System; Neuropeptides; Neurotransmitters; Pancreatic Diseases; Peripheral Nerves
Abbreviations
CGRP: calcitonin gene-related peptide; ED: embryonic day; GRP: gastrin releasing peptide; NGF: nerve growth factor; NK-1: neurokinin-1; NKA: neurokinin-A; NO: nitric oxide; NPK: neuropeptide K; NPγ: neuropeptide γ; NPY: neuropeptide Y; PACAP: pituitary adenylate cyclase activating peptide; PD: post natal day; PHI: peptide histidine isoleucine; PHM: peptide histidine methionine; SP: substance P; SP/TK: substance P/tachykinins; VIP: vasoactive intestinal polypeptide
General Concepts on Entero-Pancreatic Innervation
The innervation of the digestive tract, which is known to control a wide array of functions including smooth muscle contractility, splanchnic microcirculation, epithelial cell absorption/secretion and endocrine secretion, is composed of an intrinsic component consisting of neuronal cell bodies and nerve processes of the enteric neurons (i.e., the enteric nervous system) and an extrinsic component (i.e., nerve fibers and cellular bodies localized outside the digestive tract) [1, 2]. The extrinsic nerve fibers can be anatomically and functionally classified in afferents, conveying sensory information from the digestive system to the central nervous system, and efferent nerves, through which the central nervous system-mediated response (excitatory or inhibitory) occurs. The extrinsic fibers belong to the sympathetic and parasympathetic systems [1, 2, 3, 4].
The mammalian pancreas, like the gastrointestinal tract, embryologically develops from an outgrowth of the primitive foregut and is richly innervated being composed of a variety of myelinated or unmyelinated nerve fibers, thick nerve bundles and aggregates of neural cell bodies known as intrapancreatic ganglia. These ganglionic structures are randomly scattered throughout the pancreatic parenchyma and represent the intrinsic neural component of the pancreatic nerve supply [5, 6]. The two main extrinsic components are anatomically identified in the vagus nerves (anterior and posterior branches) and the splanchnic nerve trunks. The vagus nerves reach the pancreas directly or, alternatively, they pass across the preaortic chain of the sympathetic ganglia. Post-ganglionic sympathetic fibers, whose neural cell bodies are located in the superior mesenteric and celiac ganglia, run with the splanchnic nerves. The afferent system, primarily involved in sensory/pain transmission to the central nervous system, is composed of thin unmyelinated fibers running with either the parasympathetic pathways (vagi) or the sympathetic inputs (splanchnic nerves). The cell bodies of these nerve processes can be located either in the dorsal root ganglia (the so-called spinal afferents) or in the nodose ganglia (vagal afferents) [4, 5, 6].
Neurochemical Features of the Pancreatic Innervation
One of the major features of both enteric and pancreatic neurons is the ability to synthesize, store and release a large number of biologically active substances, including peptides, amines (5-hydroxytriptamine, noradrenaline) and other molecules such as nitric oxide (NO). A list of transmitters identified in the mammalian pancreas is reported in Table 1.
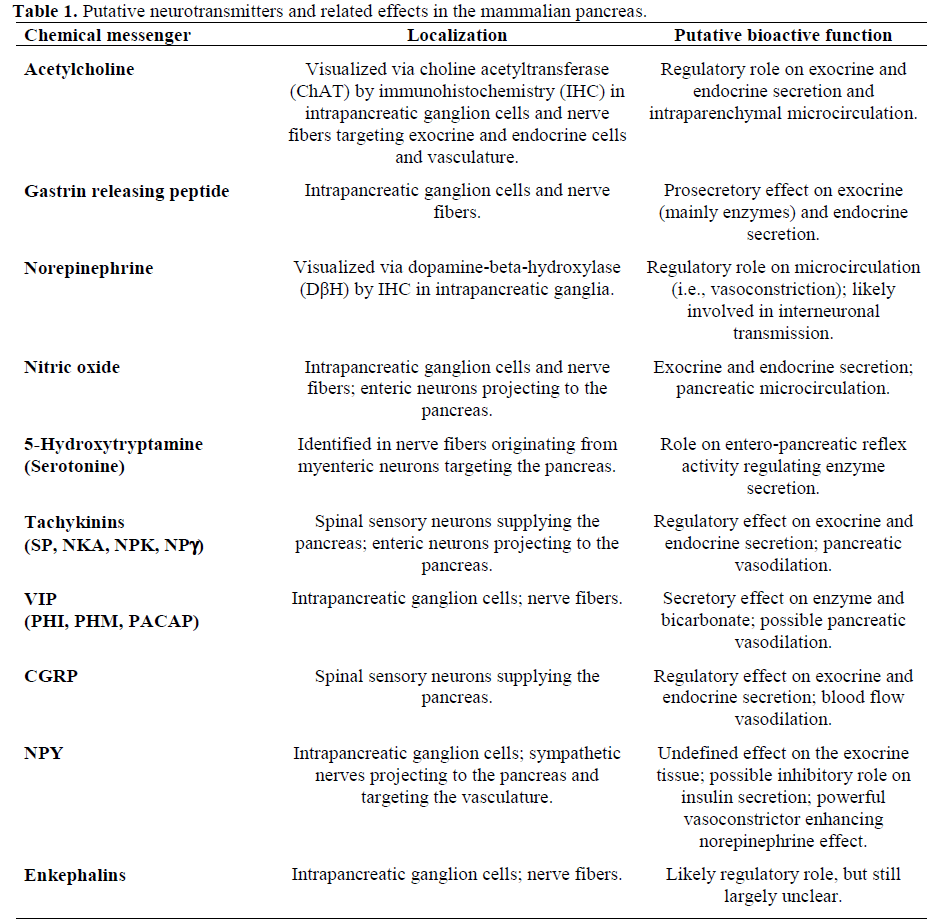
The neurochemical content, that is, the complex combination of one or more of these substances (peptides, amines, etc.) contained in the neurons is referred to as chemical coding. Over the past 20 years, the combined use of biochemical, pharmacological, electrophysiological and receptor binding methods has allowed us to recognize, identify and understand the mechanism of action of various transmitters. The surprising development of immunological techniques, mainly based on the use of specific antibodies applied to immunohistochemical techniques, has allowed the detection of distinct subpopulations of pancreatic ganglion cells and fibers depending upon their chemical content. This variety of neuronal subpopulations, identified by their neurochemistry, is known as phenotypic diversity [1, 2, 3, 4].
To better understand the neurochemistry and the regulatory mechanisms exerted by enteric and pancreatic neurons, it was necessary to analyze their development in relation to their neurotransmitters/neuromodulators expression.
Ontogenesis of the Peptide-Containing Neurons in the Gut and Pancreas
The following data are derived from studies aimed at characterizing enteric and pancreatic neurons through the identification of the expression of some neurochemical messengers, particularly neuropeptides, during development. The evidence of bioactive molecules in the enteric nervous system and pancreatic innervation of experimental animals during development contributes to a better understanding of the role played by regulatory neuropeptides in tissue differentiation and maintenance [7, 8]. The analysis of neuropeptide expression in the embryonic rat digestive system showed that among the peptides that have been studied, neuropeptide Y (NPY) is the first to appear in nerve fibers and neural cells of the gastroenteropancreatic tract. Specifically, NPY could be immunohistochemically identified in some neurons of the outermost region of the gastric mesenchyma at embryonic day (ED) 13. At ED 14, NPY immunoreactivity was detected in gastric and gut segments being localized in neural cells of the presumptive myenteric plexus (Figure 1). At ED 18, NPY was visualized in nerve processes running in the muscular layer, and in nerve fibers and cells of the submucosal plexus along the whole gastrointestinal tract. In the mucosa, NPY containing varicose fibers was observed starting from ED 20.
Figure 1. NPY immunostaining identified in neural
cells (arrows) distributed in the mesenchyma of the rat
stomach at ED 14. The NPY-containing neurons are
localized in the presumptive myenteric plexus region.
In this experiment, a rabbit polyclonal NPY antibody
(8711; kindly supplied by Prof. JH Walsh and Mrs. HE
Wong. UCLA School of Medicine. Los Angeles, CA,
USA) was applied to cryostat sections and processed
with an avidin biotin peroxidase immunohistochemical
technique. Magnification 100x.
Calcitonin gene-related peptide (CGRP) immunolabeling was detected at ED 16 in fibers and neurons of the presumptive myenteric plexus of the gut and in nerves of the gastric myenteric plexus. At ED 18, CGRP immunoreactivity was identified in fibers running through the mucosa, the submucosa, the muscular layer and around the submucosal plexus. Finally, galanin was demonstrated to appear at ED 18. This peptide was observed in nerve fibers and in neuronal cell bodies of the myenteric plexus of the gut. From ED 19 to ED 21, there is a progressive rise of the number of galanincontaining neurons both in the myenteric plexus and muscular layer along the entire digestive tract. At ED 20, galanin-containing neurons appeared in the submucosa and nerve fibers can be observed in the gut mucosa, predominantly supplying the villi. Generally, galanin innervation to the stomach and small bowel was denser than that of the colon [8].
In the pancreas, NPY and CGRP (Figure 2) appeared at ED 16 in nerves closely located to blood vessels. On that day, some nerve fibers which contained NPY were present in ducts and acini. At ED 17, CGRP fibers were found throughout the pancreas, particularly around acini and blood vessels. NPY and CGRP nerves were identified in the endocrine component of the pancreas (i.e., islets of Langerhans). At ED 18, NPY containing neural cell bodies were visualized in intrapancreatic ganglia. In contrast, CGRP immunoreactivity was present only in fibers, but not in cell bodies, of intrapancreatic ganglia. The appearance of galanin immunostaining appeared later than the other two peptides and was more evident in the postnatal period (i.e., post natal day (PD) 1) specifically being identified in nerve fibers supplying blood vessels. Subsequently, at PD 3 and PD 4, nerve fibers could be visualized in association with other pancreatic targets, such as acinar cells, ducts and intrapancreatic ganglia. In islets, galanin innervation was scanty [8].
Figure 2. CGRP immunoreactive varicose nerve
processes (arrows) supplying a blood vessel (b.v.) of
the rat pancreas at ED 16. In this experiment, a rabbit
polyclonal CGRP antibody (2p; generated by Prof. C
Sternini. UCLA School of Medicine. Los Angeles, CA,
USA) was applied to cryostat sections and processed
with an avidin biotin peroxidase immunohistochemical
technique. Magnification 100x.
Adult patterns began to be more evident in different periods of postnatal life. Hence, in the alimentary tract, NPY and galanin immunoreactivities reached adult patterns at birth (PD 0) whereas CGRP innervation reached the adult pattern at PD 2. In contrast, in the pancreas, CGRP, NPY and galanin reached the adult pattern at ED 20, PD 0, and PD 12, respectively.
The results of these studies showed a different phenotypic expression of NPY, CGRP and galanin in enteric neurons of the alimentary tract and that some neuronal subpopulations of the digestive tract and pancreas exhibited different periods of appearance and maturation. These data confirm previous analysis on the evolution and growth of the enteric and pancreatic nervous system.
The interaction between some neuropeptides could influence their own expression. For example, NPY, which appeared from ED 13, could exert a regulatory effect on the expression of peptides, such as CGRP and galanin, expressed later during development. Furthermore, some neuropeptides can interact with neurotrophic factors, such as neurotrophins (nerve growth factor, brainderived neurotrophic factors and neurotrophin-3, 4/5) in order to regulate the maturation and development of various enteric and pancreatic neurons [7, 8].
Features of Pancreatic Peptidergic Innervation: Tissue Targets and Morpho- Functional Characteristics
Using immunohistochemical methods, it was possible to thoroughly analyze the distribution of nerve fibers and neurons containing peptides in relation to a variety of innervation targets, including both exocrine (acini, ducts, blood vessels) and endocrine (islets of Langerhans) targets. Several studies from our group and other groups have clearly demonstrated that peptide-containing nerves are detectable in both the exocrine and endocrine pancreas [9, 10, 11]. A detailed analysis of specific sites of innervation in mammalian species which was carried out by our group showed that nerve fibers containing CGRP, vasoactive intestinal polypeptide (VIP) (Figure 3) and NPY, running in tiny bundles or in thick nerve trunks, were observed throughout the stroma and around acini. Overall, the analysis of the density of nerve fibers showed a more abundant number of VIP, CGRP and NPY immunolabelled processes than those positive for the gastrin releasing peptide (GRP) and substance P/tachykinins (SP/TK). The vasculature was supplied by a prominent NPY-CGRP innervation and, to a lesser extent, by galanin, SP/TK and VIP containing fibers. GRP immunoreactivity was never detected in fibers around blood vessels. Around ducts, CGRP and NPY positive neurites were more abundant than those positive for SP/TK and VIP immunoreactivity whereas GRP containing nerves were never detected. In the islets, the density of peptidergic nerves showed that VIP and GRP nerves were more numerous than those positive for CGRP, NPY or SP/TK. Within the intrapancreatic ganglia, VIP (Figure 4), galanin and, to a lesser extent, NPY immunostaining was found in numerous neuronal cell bodies and nerve processes, while GRP was present in many nerves and in a few cell bodies. Finally, CGRP and SP/TK immunoreactivities were identified only in nerve fibers surrounding unstained ganglion cells. As expected, CGRP and SP/TK were extensively colocalized in pancreatic nerves, particularly those supplying the vasculature (Figure 5) [10, 11].
Figure 3. Representative picture showing a cross
section of the pig pancreas with a dense network of
VIP-containing varicose nerves distributed throughout
the acini. Immunofluorescence technique using a VIP
rabbit polyclonal antibody (7913; kindly supplied by
Prof. JH Walsh and Mrs. HE Wong. UCLA School of
Medicine. Los Angeles, CA, USA). Magnification 80x.
Figure 4. Intrapancreatic ganglia of the cat pancreas
showing several VIP immunopositive neuronal cell
bodies (arrows) and a thick nerve bundle coming from
the ganglion and projecting towards the exocrine and
endocrine component of the gland.
Immunofluorescence technique using a VIP rabbit
polyclonal antibody (7913; kindly supplied by Prof. JH
Walsh and Mrs. HE Wong. UCLA School of Medicine.
Los Angeles, CA, USA). Magnification 80x.
Figure 5. Simultaneous visualization of CGRP (green
fluorochrome) and SP/TK (red fluorochrome) in nerve
processes (arrows) targeting a blood vessel in the rat
pancreas. The double labeling experiment was
performed using rabbit polyclonal CGRP 2p and
guinea pig SP 2f (generated by Prof. C Sternini. UCLA
School of Medicine. Los Angeles, CA, USA).
Magnification 80x.
Among the several components of the pancreatic neural supply, peptide-containing (or peptidergic) innervation is one of the most relevant; it can include an intrinsic component, originating from the intrapancreatic ganglia, and an extrinsic input of fibers which can derive from both afferent and efferent neurons. The tissue distribution and identification of specific structures (e. g., acinar cells, ducts, blood vessels, islets as well as intrapancreatic ganglia) innervated by peptide-containing nerves are crucial for understanding the sites of action of different neuropeptides.
The rich innervation of CGRP, SP/TK and, particularly, NPY fibers surrounding the vasculature provides a strong morphological basis to explain the fact that these peptides play a crucial role in the control of the intrapancreatic blood stream through a direct mechanism [4, 9, 10, 11]. Moreover, CGRP, NPY and VIP, which are widely distributed along the exocrine parenchyma (acini and ducts), are able to increase or reduce the exocrine secretion of fluid, bicarbonates (especially VIP) and of enzymes (CGRP and NPY) [4, 5, 9, 10, 11]. This secretory effect is also mediated by GRP release from nervous processes surrounding the acini [4]. The presence of GRP, VIP and CGRP fibers in the pancreatic islets support the hypothesis that these peptides could control the release of numerous pancreatic hormones [9, 10, 11]. The effect of galanin is not well-defined, although pharmacological tests showed an inhibitory effect of this peptide on insulin secretion [6, 12].
Other possible sites of neuropeptide function are represented by intrapancreatic ganglia. Anatomical and electrophysiological studies showed that pancreatic ganglia represent a complex neuronal network connected to each other by nerve bundles [13] and that single neuronal elements can develop true action potentials comparable, although not strictly identical, to those of enteric neurons [14]. Physiological tests indicate that intrapancreatic ganglia could work as intraparenchymal "pacemakers" which control hormonal and exocrine secretion [9, 10, 11, 13, 15].
In addition to the classic extrinsic (sympathetic and parasympathetic) components, it is now clear that part of the pancreatic innervation has an enteric origin [16]. Recent tracing studies, using commonly employed dyes which are taken up and transported, either retrogradely or anterogradely, through the axonal flow (Fluoro Gold, True Blue, beta-Cholera Toxin, and even herpes viruses which selectively target the neural tissue) showed that myenteric neurons from the antrum and/or the upper duodenum project to the pancreas and terminate with their neuritis in close association with different tissue targets, including acini, ducts, ganglia, vasculature and islets. The neurotransmitters released from enteric neurons extending to the pancreas include 5-hydroxytryptamine, NO and SP/TK [16]. Of remarkable interest is the observation that enzymatic secretion (amylase) may be elicited by selective stimulation of enteric-pancreatic neurons [17, 18]. These data open new perspectives in terms of understanding the pathophysiology of pancreatic diseases (e. g., acute or chronic pancreatitis) and targeted therapeutic options.
Neuropathology of Pancreatic Innervation
Experimental studies in animals and tissue analysis in patients with pancreatitis have shown significant changes in peptidecontaining nerves supplying the pancreatic gland [19, 20]. In one experimental model of chronic pancreatitis (induced by the partial legation of the Wirsung duct in the cat pancreas), we observed a significant increase in the number of VIP and NPY immunoreactive nerve fibers in the tissue involved by pancreatitis [21]. CGRP- and SP/TK- containing nerves showed a slightly denser distribution than controls while GRP nerves were not affected in this model. The neuropeptides investigated showed no apparent changes within the intrapancreatic ganglia. These results indicated a different response of peptide-containing nerves to the pancreatic damage evoked by the chronic Wirsung duct ligation. The marked expression of some neuropeptide-containing nerves, mainly VIP and NPY, suggested a neuroplasticity phenomenon occurred in the pancreatic neural network as a response to a chronic noxious stimulus. These changes based on re-arrangement of peptidecontaining innervation supplying the pancreas could be interpreted as a compensatory event secondary to chronic inflammation in order to support pancreatic secretive function and/or stimulate tissue repair mechanisms.
A further significant finding observed in this experimental model of pancreatitis is the increase in CGRP- and SP/TK-containing innervation targeting the pancreatic gland. These neural changes are in line with previous data by Büchler et al. [22] in patients with chronic calcified pancreatitis. In this study, the Authors reported a marked density of CGRP and SP/TK nerve fibers organized in thick bundles mainly found in the connective tissue surrounding fibrotic acini. Both Büchler’s results and ours showed that significant abnormalities of the CGRP and SP/TK nerve processes are of interest because they open new insights in the pathophysiology of chronic pancreatitis. Indeed, the abnormalities of the two neuropeptides CGRP and SP/TK, which are known to play a role in conveying sensory (particularly nociceptive) inputs from abdominal viscera to the central nervous system, provide a morphological basis for an impaired sensory function in the context of a chronically inflamed gland. In support of this exciting possibility, a recent study from Shrikhande et al. [23] showed an increased gene expression of the preferred receptor for SP, named neurokinin-1 (NK-1), in the tissue of patients with chronic pancreatitis. Particularly, a significant correlation between NK-1 receptor mRNA levels and the intensity of pain experienced by the patients investigated was found [23]. These findings, along with the identification of NK-1 in inflammatory cells, provided a strong basis for neuroimmune interactions likely activating sensory nerve endings in chronic pancreatitis.
Furthermore, in a model of acute pancreatitis, a possible mechanism for explaining the sensory nerve abnormalities observed was suggested to depend on the production of the nerve growth factor (NGF), a neurotrophin known to exert a major role in the neuroplastic changes and peripheral sensitization [24]. Whether this morphological finding is actually correlated with an enhanced sensory transmission deserves further study.
Conclusions
In conclusion, the mammalian pancreas is provided with a complex neural supply which innervates many tissue targets. There are distinct innervation sites depending on the peptide-containing nerve system considered. Pancreatic circuits may have both an intrinsic (intrapancreatic ganglia) or an extrinsic origin, deriving from central vagal nuclei, spinal sympathetic pre-ganglionic fibers, spinal afferent fibers and also from enteric neurons. This structural complexity, along with the multiplicity of neurotransmitters/neuromodulators identified, represents a peculiarity of the pancreatic innervation whose function is essential in controlling exocrine and endocrine pancreatic secretion. The analysis of the changes occurring in pancreatic innervation may provide new insights into the pathophysiology of a wide array of diseases, e. g., acute or chronic pancreatitis, and contribute to targeted therapeutic options.
References
- Furness JB, Costa M. The Enteric Nervous System. New York, USA: Churchill Livingston, 1987.
- Furness JB, Young HM, Pompolo S, Bornstein JC, Kunze WA, McConalogue K. Plurichemical transmission and chemical coding of neurons in the digestive tract. Gastroenterology 1995; 108:554-63. [AN 95137283]
- Gershon MD, Kirchgessner AL, Wade PR. Functional anatomy of the enteric nervous system. In: Alpers DH, ed. Physiology of the Gastrointestinal Tract. Volume 1. New York: Raven Press, 1994:381- 422.
- Furness JB, Bornstein JC. The enteric nervous system and its extrinsic connections. In: Yamada T, ed. Textbook of Gastroentrology. Volume 1. New York: Lippincott, 1991:2-24.
- Tiscornia O. The neural control of exocrine and endocrine pancreas. Am J Gastroenterol 1977; 67:541- 60. [AN 78017398]
- Holst JJ. Neural regulation of pancreatic exocrine function. In: Go VLW, ed. The Pancreas: Biology, Pathobiology, and Disease. New York: Raven Press, 1993:381-402.
- Tharakan T, Kirchgessner AL, Baxi LV, Gershon MD. Appearance of neuropeptides and NADPHdiaphorase during development of the enteropancreatic innervation. Brain Res Dev Brain Res 1995; 14:26-38. [AN 95236616]
- De Giorgio R, Sternini C, Anderson K, Brecha NC, Go VLW. Ontogenesideineuroniimmunoreattivi per galanina, neuropeptide Y e calcitonin gene-related peptide neltrattodigerente. In: Barbara L, Corinaldesi R, eds. AttiVaSettimanaItalianadelleMalattie Digestive (SIMAD V). Volume 1. Bologna, Italy: Monduzzi, 1992: 589-92.
- Su HC, Bishop AE, Power RF, Hamada Y, Polak JM. Dual intrinsic and extrinsic origins of CGRP- and NPY-immunoreactive nerves of rat gut and pancreas. J Neurosci 1987; 7:2674-87. [AN 87310613]
- De Giorgio R, Sternini C, Brecha NC, Widdison AL, Karanjia ND, Reber HA, Go VLW. Patterns of innervation of vasoactive intestinal polypeptide, neuropeptide Y and gastrin-releasing peptide immunoreactive nerves in the feline pancreas. Pancreas 1992; 7:376-84. [AN 92279188]
- De Giorgio R, Sternini C, Anderson K, Brecha NC, Go VLW. Tissue distribution and innervation pattern of peptide-immunoreactivities in the rat pancreas. Peptides 1992; 13:91-8. [AN 92319719]
- Taborsky GJ Jr, Dunning BE, Havel PJ, Ahren B, Kowalyk S, Boyle MR, Verchere CB, Baskin DG, Mundinger TO. The canine sympathetic neuropeptide galanin: a neurotransmitter in pancreas, a neuromodulator in liver. HormMetab Res 1999; 31:351-4. [AN 99349596]
- Kirchgessner AL, Pintar JE. Guinea pig pancreatic ganglia: Projections, transmitter contents, and the typespecific localization of monoamine oxidase. J Comp Neurol 1991; 305:613-31. [AN 91258627]
- King BF, Love JA, Szurszewski JH. Intracellular recordings from pancreatic ganglia of the cat. J Physiol 1989; 419:379-403. [AN 90155971]
- Stagner JI, Samols E. Role of intrapancreatic ganglia in regulation of periodic insulin secretions. Am J Physiol 1985; 248:E522-30. [AN 85196437]
- Kirchgessner AL, Gershon MD. Innervation of the pancreas by neurons in the gut. J Neurosci 1990; 10:1626-42. [AN 90237889]
- Kirchgessner AL, Gershon MD. Presynaptic inhibition by serotonin of nerve-mediated secretion of pancreatic amylase. Am J Physiol 1995; 268:G339-45. [AN 95168338]
- Kirchgessner AL, Liu MT, Gershon MD. In situ identification and visualization of neurons that mediate enteric and enteropancreatic reflexes. J Comp Neurol 1996; 371:270-86. [AN 96432680]
- Keith RG, Keshavjee SH, Kerenyi NR. Neuropathology of chronic pancreatitis in humans. Can J Surg 1985; 28:207-11. [AN 85201233]
- Di Sebastiano P, Friess H, Di Mola FF, Innocenti P, Buchler MW. Mechanisms of pain in chronicpancreatitis. Ann ItalChir 2000; 71:11-6. [AN 20288327]
- De Giorgio R, Stermini C, Widdison AL, Alvarez C, Brecha NC, Reber HA, Go VLW. Differential effects of experimentally induced chronic pancreatitis on neuropeptide immunoreactivities in the feline pancreas. Pancreas 1993; 8:700-10. [AN 94077854]
- Buechler M, Weihe E, Friess H, MalfertheinerP,Bockman E, Muller S, Nohr D, Beger HG. Changes in peptidergic innervation in chronic pancreatitis. Pancreas 1992; 7:183-92. [AN 92204914]
- Shrikhande SV, Friess H, di Mola FF, Tempia- Caliera A, Conejo Garcia JR, Zhu Z, Zimmermann A, Buchler MW. NK-1 receptor gene expression is related to pain in chronic pancreatitis. Pain 2001; 91:209-17. [AN 21173491]
- Toma H, Winston J, Micci MA, Shenoy M, Pasricha PJ. Nerve growth factor expression is upregulated in the rat model of L-arginine-induced acutepancreatitis. Gastroenterology 2000; 119:1373-81. [AN 20509975]






