Keywords
Depression; Clonidine; Agomelatine; Curcumin; Neurotransmitters
Introduction
As defined by the American psychiatric association, depression is a heterogeneous disorder associated with psychological, behavioural and physiological symptoms [1]. It occurs in all ages, genders and in all social backgrounds, it also occurs in animals. According to the World Health Organization, depression is the leading cause of disability and the 4th leading contributor to the global burden of disease with tendency to rise up to 2020 [2]. Major depression is a complex disease; it originates from the interaction between a genetic background of susceptibility and some environmental factors as stress. It is also characterized by dysfunctions of multiple systems [3] including neurotransmitters in which serotonin and norepinephrine are deficient [4], hormones, signalling pathways, neurotrophic and neuroplastic molecules and even inflammatory mediators. According to the neuroinflammatory or macrophage/cytokine theory; increased production of inflammatory cytokine causes depression via affecting a number of neural pathways including those involved with mood [5].
Animal models of depression are widely used to study the antidepressant-like effects in rodents [6]. Clonidine is α2- adrenergic receptor agonist and is widely used as anti-hypertensive agent. It produces behavioural depressive effects in laboratory animals including hypo-activity, loss of interest and investment in the environment both acutely and chronically [7]. It increases the sympatho-inhibitory function of central sympathetic neurons that leads to decrease in norepinephrine release and reduction of sympathetic tone [8].
Agomelatine is a melatonergic antidepressant (agonist to MT1 and MT2 receptors) with a rapid onset of action; it is one of the recent drugs in the antidepressant category with lower side effects than fluoxetine [9]. Melatonin is a hormone with free radical scavenging and antioxidant properties and shows its effect through MT1 and MT2 receptors [10]. It also plays an important role in the various neuropeptides and neurohormones that affect the immune system [11]. Administration of melatonin in rats increases GABA levels in the cerebellum and cerebral cortex [12]. Agomelatine also act as 5-HT2C antagonist [13]. It is well known that, 5HT2C antagonism is beneficial for alleviating different symptoms associated with 5HT2C overstimulation, including insomnia, anxiety and sexual dysfunction. Thus, the additive or perhaps even the synergistic action of agomelatine at both the melatonin and 5HT2C receptors appears to be necessary for its efficacy in treating depression [14].
Curcumin is an active ingredient in Curcuma longa Linn, more commonly known as the Asian yellow spice Turmeric [15]. Curcumin has been demonstrated to be a multi-target natural compound, which may modulate numerous pathways [16] either directly by influencing numerous signalling molecules or indirectly by modulating gene expression [17]. Curcumin has been shown to have anti-oxidant and anti-inflammatory [18], immune-modulatory [19], anti-cancer [20] and neuroprotective activities [21]. The antidepressant activity of curcumin has been explored in various animal models of depression such as forced swimming test and chronic stress model [22, 23]. Curcumin is widely consumed by the general human population and is also being employed as therapeutic agents to alleviate various neurological disorders [24].
Thus, the aim of the current study was to investigate the effect of agomelatine and/or curcumin on the behaviour, brain neurotransmitters, anti-oxidant and anti-inflammatory markers using clonidine-induced depression model in rat as well as to compare their activities with the standard antidepressant fluoxetine.
Materials and Methods
Animals
The study was performed in accordance with the ethical guidelines of Faculty of Pharmacy, Al-Azhar University, Egypt. Sixty Adult male Sprague-Dawley rats, weighing 200-220 gms were utilized. Rats were obtained from the breeding colony of the National Organization for Drug Control and Research, Giza, Egypt. They were kept under the same adequate environmental conditions at a temperature of 25 ± 1°C with alternatively 12 hour light and dark cycles and provided with their daily dietary requirements consisting of standard diet pellets (El-Nasr Chemical Co., Abu Zaabal, Cairo, Egypt) contained not less than 20% protein, 5% fibre, 3.5% fat, 6.5% ash and a vitamin mixture, water was given ad-libitum. Rats were housed in stainless-steel cages (three to four per cage) and kept at the animal house for an acclimation period of one week prior testing. Experiments were usually carried out at a fixed time around 9 AM: 3 PM.
Drugs and chemicals
Clonidine hydrochloride, fluoxetine hydrochloride and curcumin were purchased from Sigma-Aldrich Chemical Co. (St.Louis, MO, USA). Agomelatine was obtained from SEDICO Co. for Pharmaceuticals Industries, Egypt. All test substances were freshly dissolved in saline except agomelatine and curcumin were prepared in 50% DMSO/Saline.
Experimental design
Randomly chosen male rats were divided into six groups (10 rats/ each). In all groups except normal control one, depression was induced by administration of clonidine (0.8 mg/kg I.P) [7] from 8th to 14th day of treatment. All rats treated daily for 14 days as following; Group 1: Normal control (received saline only), Group 2: Depressed control group (received saline) Group 3: Depressed standard treated group (received fluoxetine 20 mg/Kg P.O) [25], Group 4: Depressed agomelatine treated group (received agomelatine 40 mg/Kg P.O) [26], Group 5: Depressed curcumin treated group (received curcumin 100 mg/Kg P.O) [27], Group 6: depressed combination or mixture-treated group (received both agomelatine & curcumin).
Rats were taken to the test situation one hour before each experiment for adaptation before examination of the testing parameters and after removing food and water from the home cage. Behavioural experiments were performed after the last injection; Open-Field Test (OFT) at day one then Forced Swimming Test (FST) at the third day. At the fifth day (19th day from the beginning of injection), rats were sacrificed by decapitation and brain was carefully removed, blotted and chilled. Neurotransmitters content (5-HT, NE and DA), oxidative stress biomarkers (MDA and GSH) and cytokines concentration (TNF-α and IL-6) were measured in brain tissue after homogenization.
Behavioural experiments
Open-Field Test (OFT): It represents a mild stressful condition [28], it is a general test for motor activity, excitability, emotionality and exploratory behaviour in rodents [29]. It consists of a square wooden box 80 cm x 80 cm × 40cm height [30], with red sides and white floor. The floor of the field was divided by black lines into 16 equal squares 4 × 4 [31, 32]. The test was performed under white light in a quiet room. The experiment performed between 9:00 AM - 3:00 PM. Experimental animals were taken from their cages alternately, placed into the central squares of the openfield and videotaped for 3 min [31]. The OFT was thoroughly wiped using 10% isopropyl alcohol and dried before application of a new rat in order to obviate possible effect on its behaviour due to odour remained from previous one [33]. The behaviour of the experimental rat in the OFT was continuously recorded during the 3 min. of the observation period [31] using coded symbols for the following parameters:
Latency- Time from putting the animal at the middle of the arena until decided to move [34], measured in seconds using a stopwatch.
Ambulation frequency-Number of squares crossed by the animal [28, 32], it was scored as a total count during a 3 min. period.
Rearing frequency-Number of time the animal stood stretched on its hind limbs with and without forelimbs support [28, 30]; it was scored during a 3 min. observation period.
Grooming frequency-Number of time of face washing and scratching with the hind leg and licking of the fur and genitals by the animal [30, 35], it was scored during a 3 min. observation period.
Forced Swimming Test (FST): It is a behavioural test used frequently to evaluate the potential efficacy of prospective antidepressant drugs in rats or mice [36, 37]. The swimming pool consists of a Plexiglas cylinder of 40cm height and 22 cm diameter with a rounded lid. The tank was filled up to 25 cm with tap water thermostatically controlled to be 25°C so the rats could not support themselves by touching the bottom with their feet [38]. The test was performed under white light in a quiet room. The experiment performed between 9:00 AM-3:00 PM. The procedure for the used FST has previously been described [39]. Two swimming sessions were conducted: an initial 10-min pre-test followed 24 hours later by a 5-min test. Following both swimming sessions, the rats were removed from cylinders, left to dry, and returned to their home cages. The water in the cylinders was changed after every other trial to avoid confounding results from urine or feces. Test sessions were videotaped for later scoring for the following parameters:
Immobility time-Time in seconds during which animal remained floating passively in the water in an upright position [32].
Struggling Time-The time in seconds during which the animal attempts to jump out of the tank, attempts to climb the walls and dive into the tank [39].
Biochemical parameters
Estimation of serotonin, norepinephrine and dopamine contents: Serotonin as well as catecholamines (norepinephrine and dopamine) was determined in rat’s brain according to the method of [40] using spectro-photoflourometer RF-5000 Shimadzu, Japan. Rats were sacrificed 24 hours after the last test quickly with minimum disturbance, to avoid any substantial changes in brain amines concentrations, which may occur within a few minutes [41]. Sacrificed animals were immediately dissected for brains on an ice-cold plate. They were either subjected for analysis immediately or stored at -80°C till the time of analysis. Then brains were weighed and homogenized in icecold solution of acidified n-butanol to obtain 10% homogenate. Homogenization was performed using glass homogenizer fitted with a glass pestle [Glas-Col (099ck5424)]. Duplicate internal standard of serotonin, norepinephrine and dopamine was prepared then neurotransmitters were extracted from the tissues ending to the aqueous phase which was divided into two portions, the first was used for the determination of serotonin while the other was used for the assessment of both norepinephrine and dopamine. Serotonin derivative was estimated by reaction with Ortho-phthalaldehyde while norepinephrine and dopamine derivative was estimated by oxidation of iodine.
Estimation of brain malondialdehyde (MDA) and reduced glutathion (GSH) contents: Determination of MDA was carried out according to the previously described method [42] with a slight modification in the incubation period [43]. Determination of brain reduced glutathione (GSH) was done using a test reagent kit (Biodiagnostic, Egypt) according to the described method [44] and according to the manufacturer’s instructions. Brain MDA and GSH contents were estimated in 10% brain homogenate in cold saline that was centrifuged at 4000 rpm for 15 mins at 4°C.
Estimation of brain tumor necrosis factor-alpha (TNF-α) and interleukine-6 (IL-6) contents: Tumor Necrosis Factoralpha (TNF-α) determination was done using a test reagent kit (eBioscience co., North America) according to the manufacturer’s instructions. Interleukien-6 (IL-6) was determined using a test reagent kit (WKEA med supplies corp, China) according to the manufacturer’s instructions. Brain TNF-α and IL-6 contents were estimated in 10% brain homogenate in cold saline that was centrifuged at 4000 rpm for 15 mins at 4°C. Calculations in the tested samples were determined as pg/g. tissue from the standard curve constructed.
Statistical Analysis
Data was expressed as the mean ± SEM. Statistical analysis was carried out by one way ANOVA followed by Tukey-Kramer multiple comparisons test to calculate significance of the difference between treatments, values of P<0.05 were considered significant. Statistical analysis was done using GRAPHPAD PRISM software (version 5, San Diego, CA, USA).
Results
Performance in the open-field test
As illustrated in Figures 1i-1iv, administration of clonidine significantly increased latency time to 300.54% while it showed a significant reduction in the ambulation, rearing and grooming frequencies to 8.77%, 32.88% and 42.91% respectively of normal control value. Treatment with agomelatine, curcumin and their combination significantly decreased the latency time to 30.27%, 22.72% and 37.81% while significantly increased the ambulation frequency to 901.89%, 986.39% and 643.35% respectively as compared to the depressed control values. Rearing frequency was also increased to 232.40%, 243.13% and 268.24% respectively as compared to depressed control values. There was a significant increase in the grooming frequency of agomelatine and agomelatine/curcumin treated rats to 233% and 283% respectively as compared to depressed control rats.
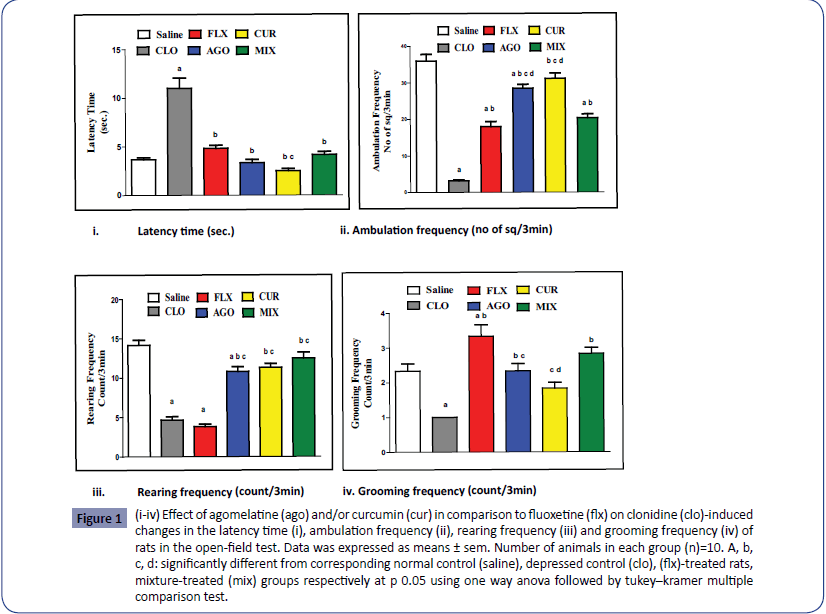
Figure 1: (i-iv) Effect of agomelatine (ago) and/or curcumin (cur) in comparison to fluoxetine (flx) on clonidine (clo)-induced changes in the latency time (i), ambulation frequency (ii), rearing frequency (iii) and grooming frequency (iv) of rats in the open-field test. Data was expressed as means ± sem. Number of animals in each group (n)=10. A, b, c, d: significantly different from corresponding normal control (saline), depressed control (clo), (flx)-treated rats, mixture-treated (mix) groups respectively at p 0.05 using one way anova followed by tukey–kramer multiple comparison test.
Performance in the forced swimming test
As shown in Figures 2i and 2ii, clonidine administration induced a significant elevation in immobility time to 209.55% while caused a significant reduction in struggling time to 23.37% as compared to the normal control rats. Treatment with agomelatine and curcumin either separately or in combination significantly decreased the immobility time to 74.67%, 58.74% and 64.93% while significantly increased the struggling time to 253.89%, 343.89% and 304.05% respectively as compared to depressed control rats.
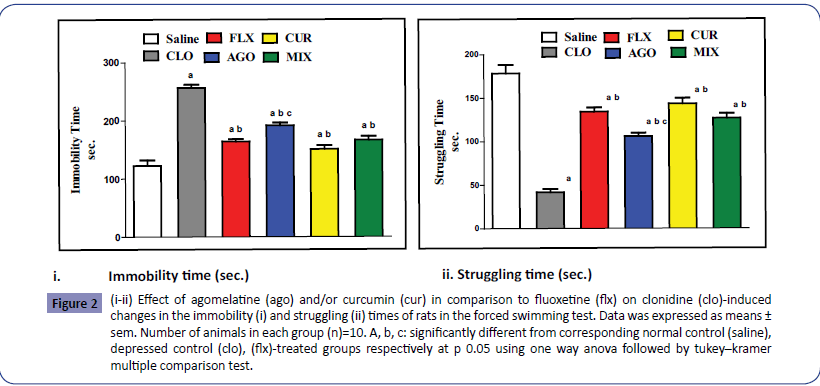
Figure 2: (i-ii) Effect of agomelatine (ago) and/or curcumin (cur) in comparison to fluoxetine (flx) on clonidine (clo)-induced changes in the immobility (i) and struggling (ii) times of rats in the forced swimming test. Data was expressed as means ± sem. Number of animals in each group (n)=10. A, b, c: significantly different from corresponding normal control (saline), depressed control (clo), (flx)-treated groups respectively at p 0.05 using one way anova followed by tukey–kramer multiple comparison test.
Brain monoamines contents
As illustrated in Figures 3i-3iii, administration of clonidine significantly decreased serotonin, norepinephrine and dopamine contents to 53.25%, 72.08% and 69.62% respectively as compared to the normal control rats. Treatment with agomelatine and curcumin either separately or in combination significantly increased serotonin content to 188.88%, 193.33% and 151.11% and norepinephrine content to 182.37%, 153.07% and 139.54% respectively of the depressed control values. Treatment with agomelatine and curcumin separately significantly increased dopamine content to 67.6% and 144.19% respectively as compared to depressed control rats while there co-administration did not show significant increase.
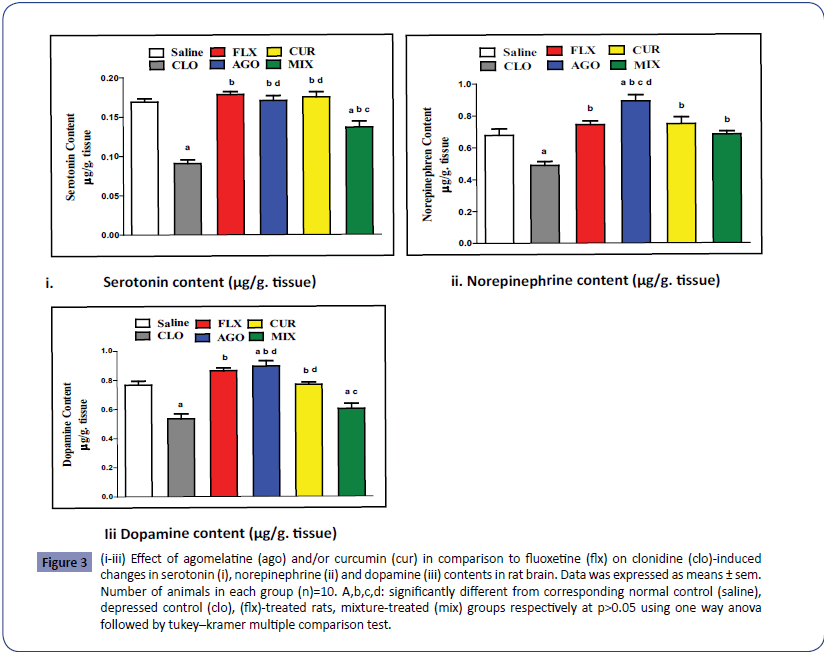
Figure 3: (i-iii) Effect of agomelatine (ago) and/or curcumin (cur) in comparison to fluoxetine (flx) on clonidine (clo)-induced changes in serotonin (i), norepinephrine (ii) and dopamine (iii) contents in rat brain. Data was expressed as means ± sem. Number of animals in each group (n)=10. A,b,c,d: significantly different from corresponding normal control (saline), depressed control (clo), (flx)-treated rats, mixture-treated (mix) groups respectively at p>0.05 using one way anova followed by tukey–kramer multiple comparison test.
Brain MDA and GSH contents
As illustrated in Figures 4i and 4ii, depression induced by administration of clonidine caused significant increase in brain MDA content reached to 112.24% with significant decrease in brain GSH content to 82.10% as compared to the normal control contents. Administration of agomelatine and curcumin either separately or in combination significantly decreased brain MDA content to 86.91%, 83.58% and 88.28% while significantly increased brain GSH content to 118.71%, 114.57% and 116.61% respectively as compared to depressed control contents. However co-administration of agomelatine and curcumin did not significantly alter either brain MDA content or GSH content as compared to each treatment alone.
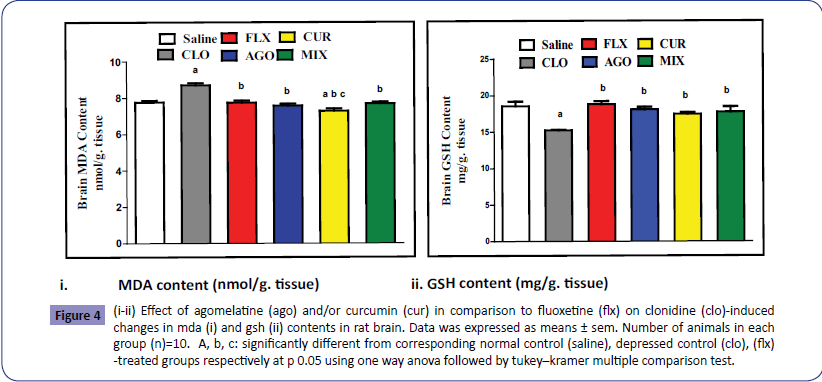
Figure 4: (i-ii) Effect of agomelatine (ago) and/or curcumin (cur) in comparison to fluoxetine (flx) on clonidine (clo)-induced changes in mda (i) and gsh (ii) contents in rat brain. Data was expressed as means ± sem. Number of animals in each group (n)=10. A, b, c: significantly different from corresponding normal control (saline), depressed control (clo), (flx) -treated groups respectively at p 0.05 using one way anova followed by tukey–kramer multiple comparison test.
Brain TNF-α and IL-6 contents
As illustrated in Figures 5i and 5ii, administration of clonidine significantly elevated brain TNF-α and brain IL-6 content to 152.32% and 137.60% respectively as compared to the normal control rats. Administration of agomelatine and curcumin either separately or in combination significantly decreased both brain TNF-α content to 51.79%, 65.43% & 71.20% and brain IL-6 content to 59.38%, 41.95% & 45.58% respectively as compared to depressed control rats. Agomelatine administration alone showed significant decrease in brain TNF-α content as compared to agomelatine and curcumin co-administration.
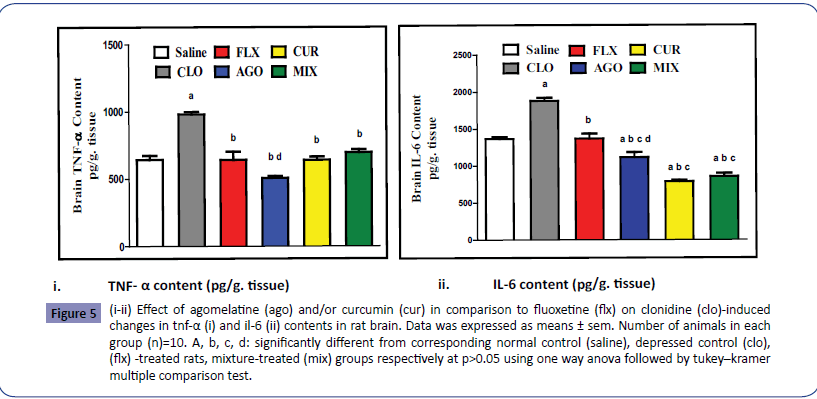
Figure 5: (i-ii) Effect of agomelatine (ago) and/or curcumin (cur) in comparison to fluoxetine (flx) on clonidine (clo)-induced changes in tnf-a (i) and il-6 (ii) contents in rat brain. Data was expressed as means ± sem. Number of animals in each group (n)=10. A, b, c, d: significantly different from corresponding normal control (saline), depressed control (clo), (flx) -treated rats, mixture-treated (mix) groups respectively at p>0.05 using one way anova followed by tukey–kramer multiple comparison test.
Discussion
Depression is an important public health issue [45] due to its high prevalence [46], the markedly reduced quality of life in both patients and their relatives [47] and the high economic burden [48]. In the present study, injection of clonidine daily for seven days resulted in depression-like behaviour in rats as indicated by significant increase in latency time as well as decrease in ambulation, rearing and grooming in the OFT while in the FST, there was an increase in immobility time with a decrease in the struggling time. These data were in accordance with the previous studies on clonidine-induced depression in rats using FST [49] or using both FST and OFT [50]. It is worthy to mention that, increased passive behaviour response in the forced swimming test such as immobility and decreased active behaviour like swimming or struggling are thought to be a clear indication of depressive like symptoms [51].
Results of the current study also showed that, clonidine induced an alteration in neurotransmitters in the form of a decrease in brain serotonin (5-HT), dopamine (DA) and norepinephrine (NE) contents. Clinical studies have revealed that the monoamines (5- HT, DA, NE) have a crucial role in the development of depression [52]. Several studies reported that 5-HT and NE have showed a drastic decrease in rats treated with clonidine (0.8 mg/kg I.P) for seven days [49, 53, 54]. The biochemical basis of clonidine– induced behavioural changes is probably due to decrease in NE release in the CNS [55]. A close inter-relation was shown to exist between NE and 5-HT [56]. It is known that clonidine administration decreased 5-HT transmission by direct activation of α2-heteroreceptors on 5-HT terminals [57]. The dopamine hypothesis indicates that the dopaminergic system strongly interacts with the norepinephrine and serotoninergic systems in the pathophysiology of depression [58].
In the present work, clonidine also produced alterations in oxidative stress biomarkers as well as pro-inflammatory cytokines in the brain of rats which was manifested by an increase in malonaldehyde (MDA), TNF-α and IL-6 contents accompanied with a decrease in reduced glutathione (GSH) content. In another research, it is reported that clonidine resulted in an increase in MDA and IL-6 accompanied with a reduction in GSH brain contents without any significant change in TNF-α [54]. Additionally, it is reported that clonidine induced significant increase in TRABS and TNF-α contents accompanied with significant depletion of GSH content [49]. Several pro-inflammatory cytokines were found to be up-regulated in depressed individuals and both oxidative and inflammatory pathways had been shown to be positively correlated with each other forming a vicious and selfenforcing cycle [59]. Reactive oxygen species were also found to activate pro-inflammatory mediators such as IL-1b, IL-6 and nuclear factor-kappa B [60]. Moreover, inflammatory mediators released by microglia in response to stress induce neurotoxicity, neurodegeneration and dysfunction of serotonergic and noradrenergic systems [61].
On the other hand, most of the currently used antidepressants have a delayed onset of action [62] as well as a variety of undesirable side effects such as sedation, decrease of blood pressure, increase of weight, indigestion or sexual dysfunction. This often results in patients’ poor compliance and break-up of medication with recurrence of depressive symptoms and increase in suicidal risk [63]. Agomelatine has been used in the current work; it is an antidepressant with rapid onset of action and relatively lower side effects, it protected rats from the behavioural alterations produced by clonidine as shown by normalized latency time and grooming frequency as well as increased ambulation and rearing frequencies in OFT. It also decreased the immobility time and increased the struggling time in FST. These data are in accordance with the previous studies concerning its antidepressant activities as well as some related stress-induced behavioural changes [64- 66]. The antidepressant-like effect of agomelatine depends on its interaction with melatonin receptors [67].
The present data also revealed that agomelatine administration reversed the decrease in monoamine (5-HT, NE, and DA) contents in the brain of depressed rats. These findings are in partial agreement with other study in which agomelatine elicited a sustained increase in dialysis levels of DA and NE in the frontal cortex of freely moving rats, whereas 5-HT was not significantly modified [68]. However, these findings are in complete agreement with another study which revealed that agomelatine through its melatonergic agonistic activity caused enhancement of DA and 5-HT neurotransmission and increased brain dopamine [26]. Bertaina-Anglade et al. [69] attributed the antidepressantlike activity of agomelatine not only to its agonistic activity at melatoninergic (MT1/MT2) receptors but also to its antagonistic activity at 5-HT2C receptors. Antagonism at 5-HT2C receptors leads to enhancement of dopaminergic and adrenergic input to the frontal cortex, induction of hippocampal neurogenesis and ultimately to agomelatine’s antidepressant effect [14].
The present data displayed that agomelatine compensated oxidative stress and inflammation caused by clonidine as demonstrated by raised brain GSH content as well as reduced brain MDA, TNF-α and IL-6 contents. These finding are parallel to those showed that agomelatine reversed the increase in TNF-α and IL-6 levels [70]. Additionally, it is reported that administration of agomelatine reversed the decrease in GSH level and the increase in TNF-α and IL-6 levels in rat’s brains [71]. Similar published findings also reported the same results except that there was no effect on IL-6 level [72]. Agomelatine protection against oxidative stress and inflammation appears to be mediated through MT1 and possibly MT2 receptors [73, 74]. Consequently, through affecting these receptors, agomelatine reset the disturbed rhythms and sleep–wake rhythms. Impaired sleep has been shown to adversely affect oxidative stress [75], mitochondrial integrity and function [76] as well as inflammation [77]; mechanisms that play pathophysiological roles in mood and anxiety disorders. Improvement of sleep by agomelatine can therefore improve oxidative, mitochondrial, and inflammatory processes which are contributed to the pathophysiology of major depression [78]. Moreover, agomelatine as an immuneregulatory agent, acts on the whole temporal progression of the inflammatory response [79] which includes an early phase characterized by the release of pro-inflammatory cytokines and a later phase that may be characterized by changes in microglia as well as in systems that may lie downstream cytokine induction [80]. Together with these results, it is suggested that the antiinflammatory effect of agomelatine involves oxidant-cleansing properties [72].
Finally, the effectiveness of the novel antidepressant agomelatine is considered to be due to its dual action of preserving sleep quality and efficiency through melatonergic MT1 and MT2 activation as well as elevating mood and activity through serotoninergic 5-HT2C antagonism [78]. It is worthy to mention that; agomelatine shows more complete profile than fluoxetine in correcting the neurochemical and behavioural abnormalities of the depressed rats [66].
Curcumin, a naturally occurring phenolic compound, is commonly used as a spice, additive and food colorant [81]. In the present study, administration of curcumin guarded rats from depression caused by clonidine injection. This effect was evidenced by the decline in latency time along with the increase in ambulation and rearing frequencies in the OFT and the decline in the immobility time as well as the rise in the struggling time in the FST. These results are in accordance with the previous data which demonstrated that curcumin significantly reversed the decrease in locomotor activity of rats induced by chronic mild stress [82]. Also, administration of curcumin to chronic unpredictable mild stress exposed rats ameliorated the decreased locomotor activities due to stress exposure as well as significantly decreased immobility duration with complementary increase in swimming time [27, 83]. According to Wang et al. [84] studies, the duration of immobility in the FST was decreased as a result of pre-treatment with curcumin in lipopolysaccharide-induced depressive-like behaviour in mice with no significant difference in the locomotor activity. Moreover, administration of curcumin simultaneously with applying chronic unpredictable stress did not affect the number of crossing and rearing of rats in OFT [85]. Curcumin has a lipophilic properties, thus it easily crosses to the blood-brain barrier or cell membranes [86, 87] and as a result possesses possible beneficial effects in experimental models of depression [88]. The antidepressant effect of curcumin might be attributed to its suppression of central 5-HT1A, 5-HT1B and 5-HT7 receptors in rats [89].
Current study showed also that curcumin administration improved central neurotransmitter (5-HT, NE, and DA) contents in the brain of the depressed rats. These results are in accordance with the study which shows that treatment with curcumin produces significant increase in 5-HT, NE and DA in the brain of depressed rats [27]. In addition, pre-treatment with curcumin showed significant improvement in the neurochemical contents in the brain of animals [90-92]. It is suggested that, the neuroprotective effects of curcumin may involve the modulation of central monoaminergic neurotransmitter systems [27]. Increased levels of serotonin, dopamine and norepinephrine by curcumin can be attributed to monoamine oxidase inhibitory effects on MAO-A and MAO-B enzymes [25, 93]. The effect of curcumin on serotonin appeared to be dose-dependent, at high doses curcumin also increased dopamine and to a lesser extent noradrenaline. Additionally, the effect of curcumin on the serotonergic system was possibly related to its interaction with 5-HT1A/1B and 5-HT2C receptors [25]. Curcumin also elevates brain DA levels and interact with D1 and D2 receptors [94].
The present data also displayed that, curcumin raised brain GSH and reduced brain MDA, TNF-α and IL-6 contents as compared to clonidine-treated rats. Besides, the reduction of brain MDA and IL-6 by curcumin was more pronounced than that of the used standard antidepressant fluoxetine. Similar results showed that chronic treatment with curcumin significantly attenuated oxidative damage as indicated by reduction in MDA and restoration of the reduced GSH levels, it also decreased the inflammation as indicated by reduction in the high level of TNF-α caused by olfactory bulbectomy in rats [88]. Along with this, curcumin effectively inhibits gene expression of these cytokine [82] and attenuates microglia activation and overproduction of TNF-α [84] as well as produces dose-dependent reduction in TNF-α levels in the brain of reserpine-depressed rats [27]. It is believed that the anti-inflammatory properties of curcumin are most likely through the dose-dependent inhibition of microglial activation and down-regulation of IL-1β, IL-6 and TNF-α in both transcriptional and translational levels [95]. Curcumin was also found to be a scavenger of the formed reactive oxygen species probably more than vitamin E and A [91]. It penetrates the inner mitochondrial membrane freely due to its lipophilic nature [24] and protects the mitochondria against various oxidative stress conditions [96, 97].
Existing data demonstrated that co-administration of agomelatine and curcumin decreased latency time with concomitant increase in ambulation, rearing and grooming frequencies in the OFT. Beyond, the immobility time was declined and struggling time was ameliorated in the FST. However, the improvement in the OFT and FST parameters were similar to either of the treatment alone except for the ambulation frequency in the OFT which was lower with both treatments while grooming was improved as compared to curcumin group. Moreover, the present study showed that the brain contents of 5-HT, NE, and DA were significantly raised in rats received combination of agomelatine and curcumin. Yet the outcome was still lower than that for each drug alone. Additionally, current investigation data revealed that brain values of oxidative stress biomarkers (MDA, GSH) and pro-inflammatory cytokines (TNF-α, IL-6) in rats given agomelatine combined with curcumin were analogue to that resulted after administration of agomelatine or curcumin separately.
Combination of curcumin with other antidepressants has shown to synergistically increase the serotonin level and enhance antidepressant-like activity in various animal models [98]. Previous researchers also detected that curcumin enhanced the antidepressant actions of escitalopram and had no significant adverse effects in depressed subjects [99]. Take into consideration that in contrast to imipramine, fluoxetine and tianeptine as well as agomelatine acts on two pharmacological substrates; melatonin and 5HT2C receptors [100]. Consequently, its application produces more distinct behavioural and molecular therapeutic profiles than that of other antidepressants.
In the present study, no additive or pronounced effects were detected by concomitant administration of curcumin and agomelatine which may be attributed to dose related reasons; for reaching the ceiling effect or due to the adaptive mechanisms occurred at the synaptic level by their repeated administration. However, there is no published data regarding the effect of the combined treatments of curcumin and agomelatine. Additionally, agomelatine’s systemic exposure increases proportionately with dose. However, at a significantly higher doses (>200 mg and above), saturation of first pass metabolism occurs, and plasma levels may increase disproportionately [101]. Clinically, drug interactions are unlikely, with the exception of co-administered potent cytochrome P1A2 (CYP1A2) enzyme inhibitors, which may increase agomelatine’s plasma level [102]. It is worthy to mention that, curcumin shows a competitive type of inhibition towards CYP1A2 [103].
On the other hand, agomelatine acts as: MT1/MT2 receptor agonist, 5-HT2C receptor antagonist [69], enhances dopaminergic and adrenergic input, induces hippocampal neurogenesis [14], decreases pro-inflammatory cytokines [70], alters microglia activity following LPS injection [80] and has antioxidant properties [72]. Meanwhile, curcumin performs its action through interaction with 5-HT1A/1B, 5-HT2C [25] and D1 and D2 receptors as well [94]. Additionally, curcumin inhibits microglial activation and down-regulates IL-1β, IL-6 and TNF-α transcription [95]. Curcumin also possess powerful antioxidant activity [91]. Consequently and in the light of what was mentioned it can be speculated that agomelatine and curcumin shared the same long-term adaptive mechanisms induced by their repeated administration, hence there was no enhanced effects in their combined administration. Further researches have been needed to explore the molecular alterations induced by agomelatine and curcumin in various models of depression in rats as well as the mechanisms of depressive-like phenotype reversion in the brain.
Conclusion
Clonidine administration causes several depressive-like symptoms and deleterious effects on neurobehavioral functions; it also induces inflammation and oxidative stress in rat’s brain. Agomelatine or curcumin in the used therapeutic doses have high efficiency against clonidine-induced abnormalities. The effect of curcumin is more pronounced than that of fluoxetine concerning the behavioural changes. However, concomitant administration of both agomelatine and curcumin against development of depression in rats has no pronounced or additive effects than each of them.
Conflict of Interest
The authors declare that they have no conflicts of interest to disclose.
Financial Support
This research has not received any specific grant from any funding agency in the public, commercial or not for profit sectors. The work has been done on the expense of the authors.
References
- Cryan JF, Markou A,Lucki I (2002) Assessing antidepressant activity in rodents: recent developments and future needs. Trends Pharmacol Sci23: 238-245.
- Lang UE,Borgwardt S (2013) Molecular mechanisms of depression: perspectives on new treatment strategies. Cell Physiol Biochem31: 761-777.
- Rossetti AC, Papp M, Gruca P, Paladini MS, Racagni G, et al. (2016) Stress-induced anhedonia is associated with the activation of the inflammatory system in the rat brain: Restorative effect of pharmacological intervention. Pharmacol Res103: 1-12.
- Raedler TJ (2011) Inflammatory mechanisms in major depressive disorder. Curr Opin Psychiatry24: 519-525.
- Maes M, Yirmyia R, Noraberg J, Brene S, Hibbeln J, et al. (2009) The inflammatory and neurodegenerative (I and ND) hypothesis of depression: leads for future research and new drug developments in depression. Metab Brain Dis24: 27-53.
- Antkiewicz-Michaluk L, Wasik A, Mozdzen E, Romanska I,Michaluk J (2014) Antidepressant-like effect of tetrahydroisoquinoline amines in the animal model of depressive disorder induced by repeated administration of a low dose of reserpine: behavioral and neurochemical studies in the rat. Neurotox Res26: 85-98.
- Enginar N, Eroglu L (1990) The long-term clonidine treatment induced behavioral depression in rats. Pol J Pharmacol Pharm42: 409-415.
- Khan ZP, Ferguson C, Jones RM (1999) Alpha-2 and imidazoline receptor agonistsTheir pharmacology and therapeutic role. Anaesthesia54: 146-165.
- Fornaro M, Prestia D, Colicchio S,Perugi G (2010) A systematic, updated review on the antidepressant agomelatine focusing on its melatonergic modulation. Curr Neuropharmacol8: 287-304.
- Brzezinski A (1997) Melatonin in humans. N Engl J Med336: 186-195.
- Reiter RJ (1995) Functional pleiotropy of the neurohormone melatonin: antioxidant protection and neuroendocrine regulation. Front Neuroendocrinol16: 383-415.
- Rosenstein RE,Cardinali DP (1986) Melatonin increases in vivo GABA accumulation in rat hypothalamus, cerebellum, cerebral cortex and pineal gland. Brain Res398: 403-406.
- Montgomery SA, Nielsen RZ, Poulsen LH,Haggstrom L (2014) A randomised, double-blind study in adults with major depressive disorder with an inadequate response to a single course of selective serotonin reuptake inhibitor or serotonin-noradrenaline reuptake inhibitor treatment switched to vortioxetine or agomelatine. Hum Psychopharmacol29: 470-482.
- de Bodinat C, Guardiola-Lemaitre B, Mocaer E, Renard P, Munoz C, et al. (2010) Agomelatine, the first melatonergic antidepressant: discovery, characterization and development. Nat Rev Drug Discov9: 628-642.
- Goel A, Jhurani S,Aggarwal BB (2008) Multi-targeted therapy by curcumin: how spicy is it? Mol Nutr Food Res52: 1010-1030.
- Zhou H, Beevers CS,Huang S (2011) The targets of curcumin. Curr Drug Targets12: 332-347.
- Gupta SC, Patchva S,Aggarwal BB (2013) Therapeutic roles of curcumin: lessons learned from clinical trials. AAPS J15: 195-218.
- Motterlini R, Foresti R, Bassi R,Green CJ (2000) Curcumin, an antioxidant and anti-inflammatory agent, induces heme oxygenase-1 and protects endothelial cells against oxidative stress. Free Radic Biol Med28: 1303-1312.
- Varalakshmi C, Ali AM, Pardhasaradhi BV, Srivastava RM, Singh S, et al. (2008) Immunomodulatory effects of curcumin: in-vivo. Int Immunopharmacol8: 688-700.
- Sharma RA, Euden SA, Platton SL, Cooke DN, Shafayat A, et al. (2004) Phase I clinical trial of oral curcumin: biomarkers of systemic activity and compliance. Clin Cancer Res10: 6847-6854.
- Xu Y, Ku B, Cui L, Li X, Barish PA, et al. (2007) Curcumin reverses impaired hippocampal neurogenesis and increases serotonin receptor 1A mRNA and brain-derived neurotrophic factor expression in chronically stressed rats. Brain Res1162: 9-18.
- Yu ZF, Kong LD,Chen Y (2002) Antidepressant activity of aqueous extracts of Curcuma longa in mice. J Ethnopharmacol83: 161-165.
- Wang R, Xu Y, Wu HL, Li YB, Li YH, et al. (2008) The antidepressant effects of curcumin in the forced swimming test involve 5-HT1 and 5-HT2 receptors. Eur J Pharmacol578: 43-50.
- Waseem M, Parvez S (2016) Neuroprotective activities of curcumin and quercetin with potential relevance to mitochondrial dysfunction induced by oxaliplatin. Protoplasma253: 417-430.
- Murad HA, Suliaman MI, Abdallah H,Abdulsattar M (2014) Does Curcumin or Pindolol Potentiate Fluoxetine's Antidepressant Effect by a Pharmacokinetic or Pharmacodynamic Interaction? Indian J Pharm Sci76: 203-210.
- Chenu F, El Mansari M,Blier P (2013) Electrophysiological effects of repeated administration of agomelatine on the dopamine, norepinephrine, and serotonin systems in the rat brain. Neuropsychopharmacology38: 275-284.
- Arora V, Kuhad A, Tiwari V,Chopra K (2011) Curcumin ameliorates reserpine-induced pain-depression dyad: behavioural, biochemical, neurochemical and molecular evidences. Psychoneuroendocrinology36: 1570-1581.
- van den Buuse M,de Jong W (1989) Differential effects of dopaminergic drugs on open-field behavior of spontaneously hypertensive rats and normotensive Wistar-Kyoto rats. J Pharmacol Exp Ther248: 1189-1196.
- Costall B, DomeneyAM, Gerrard PA, Kelly ME, NaylorRJ (1988) Zacopride: anxiolytic profile in rodent and primate models of anxiety. J Pharm Pharmacol40: 302-305.
- Cunha JM,Masur J (1978) Evaluation of psychotropic drugs with a modified open field test. Pharmacology16: 259-267.
- Vorhees CV (1974) Some behavioral effects of maternal hypervitaminosis A in rats. Teratology10: 269-273.
- Volosin M, Cancela L,Molina V (1988) Influence of adrenocorticotrophic hormone on the behaviour in the swim test of rats treated chronically with desipramine. J Pharm Pharmacol40: 74-76.
- Lazarini CA, Florio JC, Lemonica IP,Bernardi MM (2001) Effects of prenatal exposure to deltamethrin on forced swimming behavior, motor activity, and striatal dopamine levels in male and female rats. Neurotoxicol Teratol23: 665-673.
- Zbinden G (1981) Experimental methods in behavioral teratology. Arch Toxicol48: 69-88.
- Chow HL,Beck CH (1984) p-Chlorophenylalanine and p-chloroamphetamine pretreatment of apomorphine-challenged rats: effects on solitary and social behavior. Eur J Pharmacol102: 297-304.
- Porsolt RD, Le Pichon M,Jalfre M (1977) Depression: a new animal model sensitive to antidepressant treatments. Nature266: 730-732.
- Kirby LG,Lucki I (1997) Interaction between the forced swimming test and fluoxetine treatment on extracellular 5-hydroxytryptamine and 5-hydroxyindoleacetic acid in the rat. J Pharmacol Exp Ther282: 967-976.
- Porsolt RD, Anton G, Blavet N,Jalfre M (1978) Behavioural despair in rats: a new model sensitive to antidepressant treatments. Eur J Pharmacol47: 379-391.
- Einat H, Karbovski H, Korik J, Tsalah D,Belmaker RH (1999) Inositol reduces depressive-like behaviors in two different animal models of depression. Psychopharmacology (Berl)144: 158-162.
- Ciarlone AE (1978) Further modification of a fluorometric method for analyzing brain amines. Microchem J23: 9-12.
- Welch BL,Welch AS (1968) Differential activation by restraint stress of a mechanism to conserve brain catecholamines and serotonin in mice differing in excitability. Nature218: 575-577.
- Buege JA,Aust SD (1978) Microsomal lipid peroxidation. Methods Enzymol52: 302-310.
- Erdincler DS, Seven A, Inci F, Beger T,Candan G (1997) Lipid peroxidation and antioxidant status in experimental animals: effects of aging and hypercholesterolemic diet. Clin Chim Acta265: 77-84.
- Beutler E, Duron O,Kelly BM (1963) Improved method for the determination of blood glutathione. J Lab Clin Med61: 882-888.
- Mathers CD,Loncar D (2006) Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med3: e442.
- Steel Z, Marnane C, Iranpour C, Chey T, Jackson JW, et al. (2014) The global prevalence of common mental disorders: a systematic review and meta-analysis 1980-2013. Int J Epidemiol43: 476-493.
- Saarni SI, Suvisaari J, Sintonen H, Pirkola S, Koskinen S, et al. (2007) Impact of psychiatric disorders on health-related quality of life: general population survey. Br J Psychiatry190: 326-332.
- Wang PS, Simon G,Kessler RC (2003) The economic burden of depression and the cost-effectiveness of treatment. Int J Methods Psychiatr Res12: 22-33.
- El-Awdan SA, Zaki HF (2013) Gastroprotective activity of mirtazapine, escitalopram and venlafaxine in depressed rats. Afr J Pharm Pharmacol7: 2701-2709.
- Andrade S, Silveira SL, Gomez R, Barros HM,Ribeiro MF (2007) Gender differences of acute and chronic administration of dehydroepiandrosterone in rats submitted to the forced swimming test. Prog Neuropsychopharmacol Biol Psychiatry31: 613-621.
- El-Naga RN, Ahmed HI, Abd Al Haleem EN (2014) Effects of indole-3-carbinol on clonidine-induced neurotoxicity in rats: Impact on oxidative stress, inflammation, apoptosis and monoamine levels. Neurotoxicology44: 48-57.
- Mahesh R, Bhatt S, Devadoss T, Jindal A, Gautam B, et al. (2012) Antidepressant Potential of 5-HT3 Receptor Antagonist, N-n- propyl-3-ethoxyquinoxaline-2-carboxamide (6n). J Young Pharm4: 235-244.
- Kostowski W,Krzascik P (2003) Neonatal 5-hydroxytryptamine depletion induces depressive-like behavior in adult rats. Pol J Pharmacol55: 957-963.
- Zaki HF, Rizk HA (2013) Role of serotonergic and dopaminergic neurotransmission in the antidepressant effects of malt extract. Afr J Pharm Pharmacol7: 2960-2971.
- Dennis T, L'Heureux R, Carter C,Scatton B (1987) Presynaptic alpha-2 adrenoceptors play a major role in the effects of idazoxan on cortical noradrenaline release (as measured by in vivo dialysis) in the rat. J Pharmacol Exp Ther241: 642-649.
- Masse F, Hascoet M, Dailly E,Bourin M (2006) Effect of noradrenergic system on the anxiolytic-like effect of DOI (5-HT2A/2C agonists) in the four-plate test. Psychopharmacology (Berl)183: 471-481.
- Reneric JP, Bouvard M,Stinus L (2002) In the rat forced swimming test, chronic but not subacute administration of dual 5-HT/NA antidepressant treatments may produce greater effects than selective drugs. Behav Brain Res136: 521-532.
- Henry ME, Schmidt ME, Matochik JA, Stoddard EP,Potter WZ (2001) The effects of ECT on brain glucose: a pilot FDG PET study. JECT17: 33-40.
- Shelton RC, Claiborne J, Sidoryk-Wegrzynowicz M, Reddy R, Aschner M, et al. (2011) Altered expression of genes involved in inflammation and apoptosis in frontal cortex in major depression. Mol Psychiatry16: 751-762.
- Rawdin BJ, Mellon SH, Dhabhar FS, Epel ES, Puterman E, et al. (2013) Dysregulated relationship of inflammation and oxidative stress in major depression. Brain Behav Immun31: 143-152.
- Song C,Wang H (2011) Cytokines mediated inflammation and decreased neurogenesis in animal models of depression. Prog Neuropsychopharmacol Biol Psychiatry35: 760-768.
- Greden JF (2002) Unmet need: what justifies the search for a new antidepressant? J Clin Psychiatry63: 3-7.
- Keller MB, Hirschfeld RM, Demyttenaere K,Baldwin DS (2002) Optimizing outcomes in depression: focus on antidepressant compliance. Int Clin Psychopharmacol17: 265-271.
- Rainer Q, Xia L, Guilloux JP, Gabriel C, Mocaer E, et al. (2012) Beneficial behavioural and neurogenic effects of agomelatine in a model of depression/anxiety. Int J Neuropsychopharmacol15: 321-335.
- Schmelting B, Corbach-Sohle S, Kohlhause S, Schlumbohm C, Flugge G, et al. (2014) Agomelatine in the tree shrew model of depression: effects on stress-induced nocturnal hyperthermia and hormonal status. Eur Neuropsychopharmacol24: 437-447.
- Marrocco J, Reynaert ML, Gatta E, Gabriel C, Mocaer E, et al. (2014) The effects of antidepressant treatment in prenatally stressed rats support the glutamatergic hypothesis of stress-related disorders. J Neurosci34: 2015-2024.
- Papp M, Gruca P, Boyer PA,Mocaer E (2003) Effect of agomelatine in the chronic mild stress model of depression in the rat. Neuropsychopharmacology28: 694-703.
- Dekeyne A, Mannouryla Cour C, Gobert A, Brocco M, Lejeune F, et al. (2008) S32006, a novel 5-HT2C receptor antagonist displaying broad-based antidepressant and anxiolytic properties in rodent models. Psychopharmacology (Berl)199: 549-568.
- Bertaina-Anglade V, la Rochelle CD, Boyer PA,Mocaer E (2006) Antidepressant-like effects of agomelatine (S 20098) in the learned helplessness model. Behav Pharmacol17: 703-713.
- Mutlu O, Gumuslu E, Ulak G, Celikyurt IK, Akar F, et al. (2013) Antidepressant-Like Activity of Agomelatine in the Mouse Unpredictable Chronic Mild Stress Model. Drug Development Research74: 203-215.
- Karakus E, Halici Z, Albayrak A, Polat B, Bayir Y, et al. (2013) Agomelatine: an antidepressant with new potent hepatoprotective effects on paracetamol-induced liver damage in rats. Hum Exp Toxicol32: 846-857.
- Inanir S, Copoglu US, Kokacya H, Dokuyucu R, Erbas O, et al. (2015) Agomelatine Protection in an LPS-Induced Psychosis-Relevant Behavior Model. Med Sci Monit21: 3834-3839.
- Das A, McDowell M, Pava MJ, Smith JA, Reiter RJ, et al. (2010) The inhibition of apoptosis by melatonin in VSC4.1 motoneurons exposed to oxidative stress, glutamate excitotoxicity, or TNF-alpha toxicity involves membrane melatonin receptors. J Pineal Res48: 157-169.
- Choi SI, Dadakhujaev S, Ryu H, Im Kim T,Kim EK (2011) Melatonin protects against oxidative stress in granular corneal dystrophy type 2 corneal fibroblasts by mechanisms that involve membrane melatonin receptors. J Pineal Res51: 94-103.
- Kumar A,Singh A (2007) Protective effect of St. John's wort (Hypericum perforatum) extract on 72-hour sleep deprivation-induced anxiety-like behavior and oxidative damage in mice. Planta Med73: 1358-1364.
- Cirelli C,Tononi G (2004) Uncoupling proteins and sleep deprivation. Arch Ital Biol142: 541-549.
- Cirelli C, Faraguna U,Tononi G (2006) Changes in brain gene expression after long-term sleep deprivation. J Neurochem98: 1632-1645.
- Srinivasan V, Zakaria R, Othman Z, Lauterbach EC,Acuna-Castroviejo D (2012) Agomelatine in depressive disorders: its novel mechanisms of action. J Neuropsychiatry Clin Neurosci24: 290-308.
- Dantzer R, O'Connor JC, Freund GG, Johnson RW,Kelley KW (2008) From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci9: 46-56.
- Molteni R, Macchi F, Zecchillo C, Dell'agli M, Colombo E, et al. (2013) Modulation of the inflammatory response in rats chronically treated with the antidepressant agomelatine. Eur Neuropsychopharmacol23: 1645-1655.
- Gonzalez-Salazar A, Molina-Jijon E, Correa F, Zarco-Marquez G, Calderon-Oliver M, et al. (2011) Curcumin protects from cardiac reperfusion damage by attenuation of oxidant stress and mitochondrial dysfunction. Cardiovasc Toxicol11: 357-364.
- Jiang H, Wang Z, Wang Y, Xie K, Zhang Q, et al. (2013) Antidepressant-like effects of curcumin in chronic mild stress of rats: involvement of its anti-inflammatory action. Prog Neuropsychopharmacol Biol Psychiatry47: 33-39.
- Zhang L, Luo J, Zhang M, Yao W, Ma X, et al. (2014) Effects of curcumin on chronic, unpredictable, mild, stress-induced depressive-like behaviour and structural plasticity in the lateral amygdala of rats. Int J Neuropsychopharmacol17: 793-806.
- Wang Z, Zhang Q, Yuan L, Wang S, Liu L, et al. (2014) The effects of curcumin on depressive-like behavior in mice after lipopolysaccharide administration. Behav Brain Res274: 282-290.
- Liu D, Wang Z, Gao Z, Xie K, Zhang Q, et al. (2014) Effects of curcumin on learning and memory deficits, BDNF, and ERK protein expression in rats exposed to chronic unpredictable stress. Behav Brain Res271: 116-121.
- Mishra S,Palanivelu K (2008) The effect of curcumin (turmeric) on Alzheimer's disease: An overview. Ann Indian Acad Neurol11: 13-19.
- Ren SC, Suo QF, Du WT, Pan H, Yang MM, et al. (2010) Quercetin permeability across blood-brain barrier and its effect on the viability of U251 cells. Sichuan Da Xue Xue Bao Yi Xue Ban41: 751-754, 759.
- Rinwa P, Kumar A,Garg S (2013) Suppression of neuroinflammatory and apoptotic signaling cascade by curcumin alone and in combination with piperine in rat model of olfactory bulbectomy induced depression. PLoS One8: e61052.
- Li YC, Wang FM, Pan Y, Qiang LQ, Cheng G, et al. (2009) Antidepressant-like effects of curcumin on serotonergic receptor-coupled AC-cAMP pathway in chronic unpredictable mild stress of rats. Prog Neuropsychopharmacol Biol Psychiatry33: 435-449.
- Yang J, Song S, Li J,Liang T (2014) Neuroprotective effect of curcumin on hippocampal injury in 6-OHDA-induced Parkinson's disease rat. Pathol Res Pract210: 357-362.
- Singh S, Jamwal S,Kumar P (2015) Piperine Enhances the Protective Effect of Curcumin Against 3-NP Induced Neurotoxicity: Possible Neurotransmitters Modulation Mechanism. Neurochem Res40: 1758-1766.
- Lucetti ECP, Lucetti DL, da Silva AE, Ribeiro RBDM, Sampaio TMA, et al. (2016) Curcumin reversion of neurochemical and immunohistochemical alterations in brain ischemia is related to its antioxidant and anti-inflammatory properties. Journal of Medicinal Plants4: 20-29.
- Seo HJ, Wang SM, Han C, Lee SJ, Patkar AA, et al. (2015) Curcumin as a putative antidepressant. Expert Rev Neurother15: 269-280.
- Farahani MS, Bahramsoltani R, Farzaei MH, Abdollahi M,Rahimi R (2015) Plant-derived natural medicines for the management of depression: an overview of mechanisms of action. Rev Neurosci26: 305-321.
- Shi X, Zheng Z, Li J, Xiao Z, Qi W, et al. (2015) Curcumin inhibits Abeta-induced microglial inflammatory responses in vitro: Involvement of ERK1/2 and p38 signaling pathways. Neurosci Lett594: 105-110.
- Karuppagounder SS, Madathil SK, Pandey M, Haobam R, Rajamma U, et al. (2013) Quercetin up-regulates mitochondrial complex-I activity to protect against programmed cell death in rotenone model of Parkinson's disease in rats. Neuroscience236: 136-148
- Martinez-Morua A, Soto-Urquieta MG, Franco-Robles E, Zuniga-Trujillo I, Campos-Cervantes A, et al. (2013) Curcumin decreases oxidative stress in mitochondria isolated from liver and kidneys of high-fat diet-induced obese mice. J Asian Nat Prod Res15: 905-915.
- Yu JJ, Pei LB, Zhang Y, Wen ZY,Yang JL (2015) Chronic Supplementation of Curcumin Enhances the Efficacy of Antidepressants in Major Depressive Disorder: A Randomized, Double-Blind, Placebo-Controlled Pilot Study. J Clin Psychopharmacol35: 406-410.
- Kulkarni SK, Bhutani MK,Bishnoi M (2008) Antidepressant activity of curcumin: involvement of serotonin and dopamine system. Psychopharmacology (Berl)201: 435-442.
- Bourin M, Mocaer E,Porsolt R (2004) Antidepressant-like activity of S 20098 (agomelatine) in the forced swimming test in rodents: involvement of melatonin and serotonin receptors. J Psychiatry Neurosci29: 126-133.
- Fornaro M, Bandini F, Cestari L, Cordano C, Ogliastro C, et al. (2014) Electroretinographic modifications induced by agomelatine: a novel avenue to the understanding of the claimed antidepressant effect of the drug? Neuropsychiatr Dis Treat10: 907-914.
- Outhoff K (2012) Agomelatine: a review for general practitioners. South African Family Practice54: 181-187.