Keywords
Pancreatic Neoplasm
Abbreviations
PDAC pancreatic ductal adenocarcinoma Capan-1 5-FU2000 cloned Capan-1 cells resistant to 5-FU concentrations of 2 μg/mL in culture medium TDC test drug concentration 2D-PAGE 2D-polyacrylamide-gel electrophoresis
INTRODUCTION
Pancreatic ductal adenocarcinoma (PDAC) is a disease of the elderly, seen mainly in people older than 50 years of age [1]. In 2012, its incidence ranked 10th in the annual statistics of newly diagnosed carcinomas with 43,920 cases per annum. The median survival time of about 6 months and long-term survival of approximately 4% characterizes PDAC as a devastating disease even amongst malignant neoplasias [2, 3]. Thus the mortality rate for PDAC almost equals its incidence and it is currently the 4th leading cause of cancer deaths in the US [4]. Projections of cancerrelated death rates recently published by Rahib and colleagues even proclaim that pancreatic carcinoma will become the 2nd leading cancer killer by 2030 [5].
Several risk factors have been described for pancreatic cancer, including chronic pancreatitis, diabetes mellitus, smoking and germline mutations in genes such as BRCA2 [6]. A disease previously more common in men than in women, this trend has been changing in recent years, conceivably due to the rising number of women who smoke [7]. According to Raimondi, early diagnosis and increasing population longevity will lead to an increase in this lethal disease in the years to come [5, 8]. The clinical course of PDAC is predominantly that of a long asymptomatic progression leading to diagnosis, usually at an advanced stage of the cancer. With the neoplastic tumor mass most frequently localized in the pancreatic head (70% of tumors), followed by the body and then the tail, 80% of pancreatic cancer patients are already beyond curative therapy when diagnosed [1]. To date, surgery in the early stages of the disease is still the only chance of a cure. The aggressive and early metastatic behavior of PDAC make surgery a difficult choice, especially when one considers its retroperitoneal topography adjacent to major gastrointestinal vessels, making even curative resection the exception rather than the rule [3, 9]. Unfortunately, a curative, non-surgical option is still not in sight. Current chemotherapeutic regimes are restricted to neoadjuvant, adjuvant or palliative treatments, mostly using gemcitabine-based chemotherapy protocols. Recently, FOLFIRINOX (a combination therapy using oxaliplatin, 5-fluorouracil (5-FU), irinotecan and folinic acid) has shown significant survival benefits in patients with good performance status [10]. However, this palliative regimen is still being evaluated in ongoing clinical trials such as the CONKO-007 Study (EUDRACT Nr. 2009-014476- 21) amongst others [11-14]. Thus, it is still true to say that pyrimidine analogues such as gemcitabine or 5-FU, represent the backbone of pancreatic cancer treatment.
The chemotherapeutic restrictions based on PDAC's almost unique ability to acquire a variety of drug resistance mechanisms finally lead to the intensive research efforts of the past decades and the results have proven to be of importance in overcoming apoptosis induced by cytotoxics [15-23]. Recently, our group was able to contribute to that by demonstrating that membrane transporters are also of particular relevance in the chemoresistance of PDAC. Continuous treatment with 5-FU and/or gemcitabine induced the up-regulation of several multidrug resistance protein (MRP)-transporters, including MRP3, MRP4 and MRP5 in 5-FU-resistant Capan-1 cell clones (Capan-1 5-FU2000) when compared to the parent cell line [24]. MRP5, in particular, contributed to 5-FU resistance, as knock-down of MRP5 mRNA resulted in increased sensitivity towards 5-FU retreatment [24].
Nambaru and colleagues interestingly demonstrated similar results for MRP5 in Patu-02 cells [25] that, together with a study by our group in the same year [26], illustrated that altered transporter expression is important in overcoming the toxic effects of prolonged drug treatment.
To further elucidate the processes underlying adaptation towards pyrimidin alalogues - in particular 5-FU-, we now successfully established a model of chemoresistance using human pancreatic carcinoma cells - Capan-1. Thus, the establishment and characterization of, and alterations in, intracellular signal transduction in this unique 5-FU resistant cell line - designated Capan-1 5-FU2000 - offers supplementary insights and is the main focus of this report. To the best of our knowledge intracellular alterations in either pro- or anti-apoptotic pathways as well as inflammatory mediators in an acquired chemoresistance model of Capan-1 pancreatic carcinoma cells have not been reported so far.
The following results point out several proteins that showed differential regulation after adaptation towards 5-FU and thus might be relevant to tumor cell survival and might as well help to explain treatment failure in a clinical context
MATERIALS AND METHODS
Tissue Culture
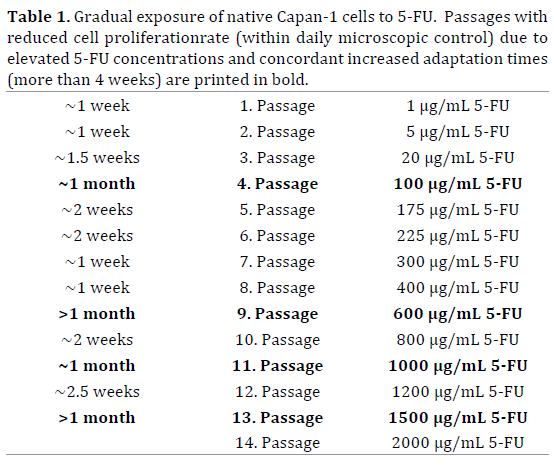
Native (parental) human pancreatic carcinoma cells (Capan-1), derived from a distant PDAC liver metastasis [27-29], were obtained from ATCC, Mannassas, VA (ATCC Number: HTB-79). The cells were adapted gradually to increasing doses of 5-FU (starting with 1 μg/mL 5-FU, added to the culture medium, and reaching 2 μg/mL within 8 months, after 14 passages) to establish acquired 5-FU-resistant Capan-1 clones (designated as Capan-1 5-FU2000).
ATP-Based Chemosensitivity Assay
The acquired resistance of the cloned Capan-1 cells was analyzed using an ATP-based assay as described previously [30-32]. Both the native Capan-1 and the cloned Capan-1 5-FU2000 cells were incubated in 96- well plates with 6 different concentrations of 5-FU for 5 to 7 days. The test drug concentration (TDC) of 5-FU was 22.5 μg/mL (172 μM = 100%) (Figure 1). All tests were performed as biological triplicates by CellTrend GmbH, Luckenwalde (Luckenwalde, Germany). Cytotoxicity was then calculated using the measured absorption rate (A) of the tested cell lines:
Cytotoxicity [%] = (1-[A treated–A blank]/[A neg–A blank]) x 100.
The inhibitory effects of 5-FU were expressed as the concentration that inhibits 50% and 90% of cell growth (IC50/IC90).
Figure 1. Chemoresistance of cloned Capan-1 5-FU2000 cells gradually
adapted to 5-FU (maximum concentration=2 μg/mL 5-FU). Incubation
time=4 days. Experiments were performed as biological triplicates by
Cell Trend GmbH, Luckenwalde (Luckenwalde, Germany).
2D-Gel Electrophoresis
Two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) was used to analyze differences in the proteomic expression patterns of native Capan-1 and Capan-1 5-FU2000 cells. Proteins were resolved by their specific isoelectric points in the first dimension, and in the second dimension by their specific molecular weight. Cells were lysed and a total of 50–75 μg of each lysate (concentration (c) = 1–2 μg protein/μL) was loaded on to linear immobilized pH gradient strips (IPG-Strip, Immobiline DryStrips pH 4–7; Amersham Biosciences, Buckinghamshire, UK). One dimensional (1D) separation was performed using the IPGphor system (Amersham Biosciences, Buckinghamshire, UK). Following 2nd dimension separation, spot detection was achieved with silver staining, or Coomassie Brilliant Blue G250 - if required for mass spectrometry. Each experiment was repeated 7 times. Subsequent spot analysis after staining (including densitometry) was performed using a GS-800 calibrated densitometer (Bio-Rad, München, Germany) and the results were analyzed using PDQuest 7.1 software (Bio-Rad, München, Germany) [33].
Quantitative Real-Time PCR (QRT-PCR)
Two-step QRT-PCR was used to quantify altered gene expression. Total RNA was isolated from cells, reversetranscribed and amplified in an Mx3000P cycler (Stratagene, Amsterdam, The Netherlands) using Absolute™ QPCR SYBR® Green Mix (Abgene, Epsom, UK). Relative quantification compared to the housekeeping gene, RPL13A, was performed using the “delta-delta-Ct” method with the following primers: S100α4, 5’-CTTGCACACGCTGTTGCTAT- 3’ and 5’-AACTTGCTCAGCATCAAGCA-3’ (467 bp fragment, bases 73–539) [34].
Western Blotting
The expression of specific proteins was analyzed using Western blotting. Equal amounts of native and 5-FU-resistant Capan-1 cell lysates were separated by SDS-PAGE, blotted onto polyvinylidene difluoride (PVDF) membranes and probed using specific primary antibodies and horseradish peroxidase (HRP)-conjugated secondary antibodies. Results were visualized using the chemiluminescence SuperSignal West Femto Chemiluminescent kit, Pierce (Pierce, Rockford, IL, USA, now part of ThermoFisher Scientific). Each test was performed as biological triplicates. Testing of the proapoptotic proteins, BOK and BAD, was performed according to the manufacturer’s instructions using the following kits: Phototope®-HRP Western Blot Detection System #7071 (Cell Signaling Technology Inc., Danvers, MA revised 04/15/08), and the Pro-Apoptosis Bcl-2 Family Antibody Sampler Kit #9942 (Cell Signaling Technology Inc., revised 01/11/07). Since these experiments were performed only once in each case, the results are only indicative.
ELISA
Cells were lysed according to the manufacturer's protocol using cell lysis buffer® #980 (Cell Signaling Technology Inc., rev. 08/16/07).
Phosphosensitive semiquantitative analysis was performed for the following proteins: ribosomal protein S6 (rpS6), nuclear factor kappa B (NfkB), nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IkB alpha), p38 alpha and stress-activated protein kinase/c-Jun N-terminal kinase (SAPK/JNK) using the PathScan® Cell Growth Multi-Target Sandwich ELISA Kit #7239 (Cell Signaling Technology Inc., new 08/07), and the PathScan® Inflammation Multi-Target Sandwich ELISA Kit #7276, (Cell Signaling Technology Inc., new 10/07) according to the manufacturer's protocol.
Treatment groups are as follows:
• Parental Capan-1: no 5-FU;
• Resistant Capan-1 (abstention): withdrawal of 5-FU from the culture medium for 4 days;
• Parental Capan-1 (treatment): 2 μg/mL 5-FU for 4 days
• Resistant Capan-1 (abstention) + retreated: 5-FU withdrawal for 4 days followed by the addition of 2 μg/ mL 5-FU for a further 4 days.
The treatment duration was 4 days during which time the medium containing 5-FU was replenished every 48 hours.
The results are presented as the mean of 2 technical replicates.
RESULTS
To achieve 5-FU resistance, chemosensitive Capan-1 cells were incubated with increasing concentrations of 5-FU in the culture medium (Table 1). To confirm that the Capan-1 5-FU2000 cloned cell line was indeed resistant to 5-FU, an ATP-based chemosensitivity assay was performed by CellTrend GmbH, Luckenwalde (Luckenwalde, Germany). The resistant cell line demonstrated a 27-fold increase in tolerance towards 5-FU, when the inhibitory concentration (IC) 50 values were compared to the native Capan-1 control cells: 63.6 μg/mL 5-FU in Capan-1 5-FU2000 (concentration based on tested IC values) vs. 2.4 μg/mL in native Capan-1 cells (Figure 1). In this experiment, 2 μg/ mL 5-FU resulted in a growth inhibition in ~50% of native Capan-1 cells (IC50).
To detect intracellular changes in protein expression, a multivariate approach was undertaken: 2D-PAGE was performed followed by gel staining and characterization of a protein spot that was clearly differentially regulated between the native and resistant cell lines (Figure 2). The differentially regulated spot was characterized via mass spectrometry as S100α4, a calcium-binding protein. In addition, Western blotting was performed to validate the observed difference in the respective proteomic expression patterns. The results confirmed a marked decrease in the expression of S100α4 at the protein level in 5-FU resistant cells (Figure 3). In the current study, S100α4 expression in 5-FU-resistant Capan-1 cells was independent of 5-FU treatment as resistant cells showed a comparable downregulation of S100α4 after withdrawal of 5-FU from the culture medium.
Figure 2. 2D-PAGE showing a differentially regulated spot after silver
staining. The spot was identified subsequently as S100α4 via mass
spectrometry.
Figure 3. Western blotting showing S100α4 expression suppressed in
5-FU-resistant Capan-1 clones.
Following ELISA assays, native cells and resistant Capan-1 cells revealed comparable basal levels of NfkB p65 and IkB-alpha phosphorylation. Treatment with 5-FU induced a marked increase in the phosphorylation of NfkB p65 and IkB-alpha in native Capan-1 cells; however, in resistant cells, 5-FU treatment decreased NfkB p65 phosphorylation, but had no effect on the phosphorylation of IkB-alpha (Figures 4 and 5).
Figure 4. Effect of 5-FU treatment on NfkB p65 phosphorylation in native and 5-FU resistant cells.
Figure 5. Effect of 5-FU treatment on IkB-alpha phosphorylation in native and 5-FU resistant cells.
The basal phosphorylation of ribosomal protein S6 was much higher in 5-FU-resistant clones than in the native cell line. Retreatment with 5-FU after 4 days of abstention interestingly resulted in decreased rpS6 phosphorylation in 5-FU-resistant cells, but had the opposite effect in native Capan-1 cells in case of treatment with 5-FU (2 μg/mL) (Figure 6).
Figure 6. Effect of 5-FU treatment on ribosomal protein S6 phosphorylation in native and 5-FU resistant cells.
ELISA testing for SAPK/JNK and P38 phosphorylation in 5-FU-resistant clones revealed a decrease in the basal phosphorylation levels of these proteins compared to the native cell line (Figures 7 and 8).
Figure 7. Effect of 5-FU treatment on SAPK/JNK phosphorylation in native and 5-FU resistant cells.
Figure 8. Effect of 5-FU treatment on p38-alpha phosphorylation in native and 5-FU resistant cells.
Supplementary western blotting experiments indicated differential expression of the pro-apoptotic mediators, BOK and BAD. Compared to the native cell line, BOK was down-regulated in 5-FU-resistant Capan-1 cells cultivated in medium with 2 μg/mL 5-FU. Withdrawal of 5-FU for 4 days increased BOK expression to levels comparable to native Capan-1 cells (Figure 9, upper panel). In contrast, BAD expression was up-regulated in 5-FU-resistant Capan-1 cells, independent of stimulation with 5-FU (Figure 9, middle panel). Actin was used as the loading control (Figure 9, lower panel).
Figure 9. Differentially regulated expression levels of BOK and BAD in (1) Capan-1 native cells: medium contained no 5-FU; (2) Capan-1 5-FU2000 cells
(resistant), abstention: 5-FU withdrawn from medium; (3) Capan-1 5-FU2000 cells (resistant): medium contained regular 5-FU dose of 2 μg/mL. The blue
square (see BOK line3 represents the densitometric area measured. Duration of treatment/incubation=22 hours.
DISCUSSION
Various cellular mechanisms have been characterized in recent decades. With respect to the model of acquired chemoresistance presented in the current study, we took a further step in elucidating intracellular differences in signal transduction pathways in Capan-1 cell lines adapted to drug-resistance. Differences in expression patterns of transporter proteins in these cells, in particular the multidrug resistance protein (MRP) family, have been described previously by our group with respect to known alterations in other tumor types displaying a resistance phenotype.
In the current study, we successfully established 5-FUresistant Capan-1 cell clones (Capan-1 5FU2000) following gradual exposure of chemosensitive cells to 5-FU during culture. We thereby demonstrate that acquired 5-FU resistance is associated with alterations in the proteomic expression pattern as well as the phosphorylation levels of some of the respective pro- and anti-apoptotic, and inflammatory regulator proteins.
First of all proving the acquired 5-FU resistance of our cell clones by comparing the IC 50 values of Capan-1 5FU2000 to the native cell line we observed a 27-fold increase in tolerance to 5-FU in the resistant cell line (Capan-1 5FU2000 = 63.6 μg/mL vs. 2.4 μg/mL in native Capan-1 cells. The concentration was based on the tested IC values - Figure 1). Comparative analysis of the parent and resistant cell lines clearly revealed a differential regulation of various pro- and anti-apoptotic mediators and of proteins known to be involved in cellular inflammation. We verified a decrease in expression of the calcium binding protein, S100α4, in resistant Capan-1 5FU2000 cell lines compared to the Capan-1 parent cells. S100α4 is thought to be involved in the regulation of chemoresistance and metastasis. Since previous reports have demonstrated that S100α4 knockdown sensitizes pancreatic carcinoma cells to gemcitabine treatment, S100α4 has been tested as an early diagnostic marker in various cancers, including the pancreas [35-38]. Furthermore, it is known that S100α4 is capable of suppressing BNIP3 promoter activity via the activation of NfkB, which is associated with chemoresistance in PDAC cell lines [37, 39]. The current results agreed with our previously published data, which demonstrated the down-regulation of S100α4 expression by 82%, and confirmed this down-regulation at the RNA level in 5-FU-resistant Capan-1 cells [34]. However, in contrast to the referred data of our group and the given data from other researchers, it remains contradictory if knockdown of S100α4 is associated with an increased sensitivity of pancreatic cancer cells towards cytotoxic treatment [34-40].
Our obtained data also conclusively reinforces the potential involvement of S100α4 in acquired chemoresistance as it illustrates a direct drug-related alteration in proteomic expression besides demonstrating the interdependence of redundantly regulated inflammatory signaling pathways, such as NfkB. The proteomic alteration fits quite well with a concordant decrease in S100α4 [34] since we observed a marked down-regulation of NfkB p65 and IkB-alpha phosphorylation in Capan-1 5FU2000 clones, according to the decrease in S100α4 levels in the resistant cell line. Those changes in signal transduction pathway responsiveness are obviously established during the adaptation of the cell line in response to a permanent 5-FU stimulus. In contrast, when native Capan-1 cells were treated with 5-FU (2 μg/ mL for 96 h; Figure 4) we observed a marked increase in NfkB phosphorylation levels. This effect might, therefore, be considered as an early cellular mechanism that has to be bypassed whilst establishing survival mechanisms in response to consequent cytotoxic stress.
With respect to our results regarding the expression of SAPK/JNK and P38 (Figures 7 and 8), their decrease in basal phosphorylation levels in case of Capan-1 5FU2000 cells also fits quite well with the rest of our data. Since phosphorylation represents a physiological cellular activation in context of cellular stress [41-44] in response to e.g. osmotic-, ultraviolet- or pro-inflammatory cytokine- mediated stress this might best be assumed as cellular habituation while permanent 5-FU exposure selects for adequate signal transduction response mechanisms [41- 49]. Thus their respective signaling pathways might necessarily be bypassed in the process of establishing an acquired 5-FU resistance to circumvent apoptosis.
We moreover found that the phosphorylation of ribosomal protein S6 (rpS6) showed a marked increase in resistant clones, but a decrease in response to treatment with 5-FU. Again this was in contrast to native Capan-1 cells where rpS6 phosphorylation was regulated with respect to the 5-FU treatment (Figure 6). Ribosomal protein S6 is thought to be important in the translation of mRNA into proteins needed for cell cycle progression as well as in the process of cellular translation itself [50, 51]. While rpS6 phosphorylation is known to be linked with increased intracellular translation frequencies, this result might be indicative of cytotoxic-induced elevated translation rates due to cellular stress [50]. Once more, the observed results are in line with elevated phosphorylation levels seen in Capan-1 5FU2000 clones, particularly since the measured basal expression levels of rpS6 were equal for native and resistant cells. One possible regulator responsible for this phenomenon might be m-TOR, a serine/threonine protein kinase known to be of relevance in tumor growth mostly via its role in PI3K/AKT signaling [52].
Using western blotting the expression patterns of the pro-apoptotic mediators, BOK and BAD, were down- and up-regulated respectively in 5-FU-resistant cell clones (Figure 9) [53-56]. In the case of BOK, it again seems reasonable that pro-apoptotic protein levels decrease as a consequence of cellular adaptation towards 5-FU exposure. Nevertheless, this was not the case for BAD. The observed increase in BAD protein levels might be best interpreted either as a loss-of-function in this signaling cascade, or it is possible that the observed increase in BAD protein is balanced by an undiscovered anti-apoptotic mediator [57]. However, it should be emphasized that the western blots presented in the current study are strictly indicative since the experiments were performed just once.
In summary, we successfully established 5FU-resistant Capan-1 cell clones adapted from a chemosensitive native Capan-1 cell line. Comaparing both cell lines we revealed a differential regulation of key mediators known to be relevant for inflammatory- as well as pro- and antiapoptotic cell signaling pathways when regarding their respective expression- and phosphorylation pattern.
The elegance of this model might be better assessed in the context of a recent publication by Jones and colleagues who presented a broad-spectrum genetic analysis of 24 pancreatic cancer samples. In their study, they discovered an average of 63 genetic alterations in every case of PDAC, which affected a core of 12 overlapping signaling pathways that were each genetically modified in 67%– 100% of the analyzed samples [58]. This result illustrated that a relatively large number of genetic alterations can affect a reasonably small number of core pathways. Those findings by Jones and colleagues call on targeted therapies that address nodal points or redundant regulators in cell signaling mechanisms rather than trying to target single mutated genes. Thus the next step in pancreatic cancer treatment becomes apparent and may allow us to kill two birds with one stone in helping to address the modern problem of cellular drug resistance. However, as we know from other types of cancer, and have seen in the current study, the processes underlying the hallmarks of cancer are mostly regulated by redundant pathways, thus the targeted treatment of only one cancer cell pathway may be insufficient as it will enable cancer cell survival following the activation of redundant alternative pathways culminating in drug resistance [59, 60].
CONCLUSION
In the current study, we successfully established 5-FUresistant Capan-1 cell clones (Capan-1 5FU2000) following gradual exposure of chemosensitive cells to 5-FU during culture. We thereby demonstrate that acquired 5-FU resistance is associated with alterations in the proteomic expression pattern as well as the phosphorylation levels of some of the respective pro- and anti-apoptotic, and inflammatory regulator proteins. Our model might therefore represent a paradigm to discover and identify starting points and potential targets for future therapies that aim at overcoming the difficulty of acquired chemoresistance.
Acknowledgments
This work was supported by a grant from the Ministry for Science, Research and the Arts (MWK), Stuttgart, Germany; by Dr. Hans & Lore Graf-Stiftung, Kaiserslautern, Germany, and by the National Swedish Science Foundation (VR K2008-55X-20655-01-3).
Authors’ Contribution
Marvin Schober: Literature research, manuscript writing; experiments
Ralf Jesenofsky, Ralf Faissner, Wolfgang Hagmann: supervision, manuscript writing
Sebastian Krug, Cornelius Weidenauer: manuscript review and writing
Rainer L Heuchel, Stephan L Haas: manuscript revision and writing
J Matthias Löhr: study design, idea, supervision, manuscript review and writing. All authors have read and approved the final version of the manuscript.
Conflicting Interest
The authors had no conflicts of interest
References
- Hidalgo M. Pancreatic cancer. The New England journal of medicine 2010; 362:1605-17. [PMID: 20427809].
- Kleeff J, Michalski C, Friess H, Buchler MW. Pancreatic cancer: From bench to 5-year survival. Pancreas 2006; 33:111-8. [PMID: 16868475].
- Löhr M. Is it possible to survive pancreatic cancer? Nature clinical practice. Gastroenterology & hepatology 2006; 3:236-7. [PMID: 16672986].
- Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA: a cancer journal for clinicians 2012; 62:10-29. [PMID: 22237781].
- Rahib L, Smith BD, Aizenberg R, Rosenzweig AB, Fleshman JM, Matrisian LM. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the united states. Cancer research 2014; 74:2913-21. [PMID: 24840647].
- Lowenfels AB, Maisonneuve P. Risk factors for pancreatic cancer. Journal of cellular biochemistry 2005; 95:649-56. [PMID: 15849724].
- Raimondi S, Maisonneuve P, Löhr JM, Lowenfels AB. Early onset pancreatic cancer: Evidence of a major role for smoking and genetic factors. Cancer epidemiology, biomarkers & prevention : a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology 2007; 16:1894-7. [PMID: 17855711].
- Raimondi S, Maisonneuve P, Lowenfels AB. Epidemiology of pancreatic cancer: An overview. Nature reviews. Gastroenterology & hepatology 2009; 6:699-708. [PMID: 19806144].
- Lüttges J, Kloppel G. [pancreatic ductal adenocarcinoma and its precursors]. Der Pathologe 2005; 26:12-7. [PMID: 15630571].
- Conroy T, Desseigne F, Ychou M, Bouche O, Guimbaud R, Becouarn Y, Adenis A, et al. Folfirinox versus gemcitabine for metastatic pancreatic cancer. The New England journal of medicine 2011; 364:1817-25. [PMID: 21561347].
- Hwang IG, Jang JS, Oh SY, Lee S, Kwon HC, Lee GW, Go S, et al. A phase ii trial of erlotinib in combination with gemcitabine and cisplatin in advanced pancreatic cancer. Investigational new drugs 2012. [PMID: 22302349].
- Lim KH, Chung E, Khan A, Cao D, Linehan D, Ben-Josef E, Wang-Gillam A. Neoadjuvant therapy of pancreatic cancer: The emerging paradigm? The oncologist 2012. [PMID: 22250057].
- Reni M, Balzano G, Aprile G, Cereda S, Passoni P, Zerbi A, Tronconi MC, et al. Adjuvant pefg (cisplatin, epirubicin, 5-fluorouracil, gemcitabine) or gemcitabine followed by chemoradiation in pancreatic cancer: A randomized phase ii trial. Annals of surgical oncology 2012. [PMID: 22237835].
- Katz MH, Fleming JB, Lee JE, Pisters PW. Current status of adjuvant therapy for pancreatic cancer. The oncologist 2010; 15:1205-13. [PMID: 21045189].
- Borst P, Schinkel AH, Smit JJ, Wagenaar E, Van Deemter L, Smith AJ, Eijdems EW, et al. Classical and novel forms of multidrug resistance and the physiological functions of p-glycoproteins in mammals. Pharmacology & therapeutics 1993; 60:289-99. [PMID: 7912835].
- Burger H, Nooter K, Zaman GJ, Sonneveld P, van Wingerden KE, Oostrum RG, Stoter G. Expression of the multidrug resistance-associated protein (mrp) in acute and chronic leukemias. Leukemia 1994; 8:990-7. [PMID: 7911548].
- Grant CE, Valdimarsson G, Hipfner DR, Almquist KC, Cole SP, Deeley RG. Overexpression of multidrug resistance-associated protein (mrp) increases resistance to natural product drugs. Cancer research 1994; 54:357-61. [PMID: 8275468].
- Bell DR, Gerlach JH, Kartner N, Buick RN, Ling V. Detection of p-glycoprotein in ovarian cancer: A molecular marker associated with multidrug resistance. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 1985; 3:311-5. [PMID: 2857774].
- Kartner N, Riordan JR, Ling V. Cell surface p-glycoprotein associated with multidrug resistance in mammalian cell lines. Science 1983; 221:1285-8. [PMID: 6137059].
- Kartner N, Shales M, Riordan JR, Ling V. Daunorubicin-resistant chinese hamster ovary cells expressing multidrug resistance and a cell-surface p-glycoprotein. Cancer research 1983; 43:4413-9. [PMID: 6135505].
- Ling V, Kartner N, Sudo T, Siminovitch L, Riordan JR. Multidrug-resistance phenotype in chinese hamster ovary cells. Cancer treatment reports 1983; 67:869-74. [PMID: 6354434].
- Riordan JR, Deuchars K, Kartner N, Alon N, Trent J, Ling V. Amplification of p-glycoprotein genes in multidrug-resistant mammalian cell lines. Nature 1985; 316:817-9. [PMID: 2863759].
- Schober M, Jesenofsky R, Faissner R, Weidenauer C, Hagmann W, Michl P, Heuchel RL, et al. Desmoplasia and chemoresistance in pancreatic cancer. Cancers (Basel) 2014; 6:2137-54. [PMID: 25337831].
- Hagmann W, Jesnowski R, Faissner R, Guo C, Lohr JM. Atp-binding cassette c transporters in human pancreatic carcinoma cell lines. Upregulation in 5-fluorouracil-resistant cells. Pancreatology 2009; 9:136-44. [PMID: 19077464].
- Nambaru PK, Hubner T, Kock K, Mews S, Grube M, Payen L, Guitton J, et al. Drug efflux transporter multidrug resistance-associated protein 5 affects sensitivity of pancreatic cancer cell lines to the nucleoside anticancer drug 5-fluorouracil. Drug Metab Dispos 2010; 39:132-9. [PMID: 20930123].
- Hagmann W, Jesenofsky R, Lohr JM. Interdependence of gemcitabine treatment, transporter expression, and resistance in human pancreatic carcinoma cells. Neoplasia 2010; 12:740-7. [PMID: 20824050].
- Moore PS, Sipos B, Orlandini S, Sorio C, Real FX, Lemoine NR, Gress T, et al. Genetic profile of 22 pancreatic carcinoma cell lines. Analysis of k-ras, p53, p16 and dpc4/smad4. Virchows Archiv : an international journal of pathology 2001; 439:798-802. [PMID: 11787853].
- Sipos B, Moser S, Kalthoff H, Torok V, Lohr M, Kloppel G. A comprehensive characterization of pancreatic ductal carcinoma cell lines: Towards the establishment of an in vitro research platform. Virchows Archiv : an international journal of pathology 2003; 442:444-52. [PMID: 12692724].
- Kyriazis AP, Kyriazis AA, Scarpelli DG, Fogh J, Rao MS, Lepera R. Human pancreatic adenocarcinoma line capan-1 in tissue culture and the nude mouse: Morphologic, biologic, and biochemical characteristics. The American journal of pathology 1982; 106:250-60. [PMID: 6278935].
- Andreotti PE, Cree IA, Kurbacher CM, Hartmann DM, Linder D, Harel G, Gleiberman I, et al. Chemosensitivity testing of human tumors using a microplate adenosine triphosphate luminescence assay: Clinical correlation for cisplatin resistance of ovarian carcinoma. Cancer research 1995; 55:5276-82. [PMID: 7585588].
- Kurbacher CM, Cree IA. Chemosensitivity testing using microplate adenosine triphosphate- based luminescence measurements. Methods in molecular medicine 2005; 110:101-20. [PMID: 15901931].
- Hunter EM, Sutherland LA, Cree IA, Dewar JA, Preece PE, Wood RA, Linder D, et al. Heterogeneity of chemosensitivity in human breast carcinoma: Use of an adenosine triphosphate (atp) chemiluminescence assay. European journal of surgical oncology : the journal of the European Society of Surgical Oncology and the British Association of Surgical Oncology 1993; 19:242-9. [PMID: 8314382].
- Lohr JM, Faissner R, Koczan D, Bewerunge P, Bassi C, Brors B, Eils R, et al. Autoantibodies against the exocrine pancreas in autoimmune pancreatitis: Gene and protein expression profiling and immunoassays identify pancreatic enzymes as a major target of the inflammatory process. The American journal of gastroenterology 2010; 105:2060-71. [PMID: 20407433].
- Hagmann W, Faissner R, Schnölzer M, Löhr JM, Jesnowski R. Membrane drug transporters and chemoresistance in human pancreatic carcinoma. Cancers 2011; 3:106-125. [PMID: 24212609].
- Kikuchi N, Horiuchi A, Osada R, Imai T, Wang C, Chen X, Konishi I. Nuclear expression of s100a4 is associated with aggressive behavior of epithelial ovarian carcinoma: An important autocrine/paracrine factor in tumor progression. Cancer Sci 2006; 97:1061-9. [PMID: 16984379].
- Cabezon T, Celis JE, Skibshoj I, Klingelhofer J, Grigorian M, Gromov P, Rank F, et al. Expression of s100a4 by a variety of cell types present in the tumor microenvironment of human breast cancer. Int J Cancer 2007; 121:1433-44. [PMID: 17565747].
- Mahon PC, Baril P, Bhakta V, Chelala C, Caulee K, Harada T, Lemoine NR. S100a4 contributes to the suppression of bnip3 expression, chemoresistance, and inhibition of apoptosis in pancreatic cancer. Cancer research 2007; 67:6786-95. [PMID: 17638890].
- Ohuchida K, Mizumoto K, Egami T, Yamaguchi H, Fujii K, Konomi H, Nagai E, et al. S100p is an early developmental marker of pancreatic carcinogenesis. Clinical cancer research : an official journal of the American Association for Cancer Research 2006; 12:5411-6. [PMID: 17000674].
- Akada M, Crnogorac-Jurcevic T, Lattimore S, Mahon P, Lopes R, Sunamura M, Matsuno S, et al. Intrinsic chemoresistance to gemcitabine is associated with decreased expression of bnip3 in pancreatic cancer. Clinical cancer research : an official journal of the American Association for Cancer Research 2005; 11:3094-101. [PMID: 15837765].
- Fernandez-Fernandez MR, Veprintsev DB, Fersht AR. Proteins of the s100 family regulate the oligomerization of p53 tumor suppressor. Proc Natl Acad Sci U S A 2005; 102:4735-40. [PMID: 15781852].
- Kyriakis JM, Avruch J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiological reviews 2001; 81:807-69. [PMID: 11274345].
- Raman M, Chen W, Cobb MH. Differential regulation and properties of mapks. Oncogene 2007; 26:3100-12. [PMID: 17496909].
- Zarubin T, Han J. Activation and signaling of the p38 map kinase pathway. Cell research 2005; 15:11-8. [PMID: 15686620].
- Leppa S, Bohmann D. Diverse functions of jnk signaling and c-jun in stress response and apoptosis. Oncogene 1999; 18:6158-62. [PMID: 10557107].
- Raingeaud J, Gupta S, Rogers JS, Dickens M, Han J, Ulevitch RJ, Davis RJ. Pro-inflammatory cytokines and environmental stress cause p38 mitogen-activated protein kinase activation by dual phosphorylation on tyrosine and threonine. The Journal of biological chemistry 1995; 270:7420-6. [PMID: 7535770].
- Whitmarsh AJ, Davis RJ. Structural organization of map-kinase signaling modules by scaffold proteins in yeast and mammals. Trends in biochemical sciences 1998; 23:481-5. [PMID: 9868371].
- Davis RJ. Signal transduction by the c-jun n-terminal kinase. Biochemical Society symposium 1999; 64:1-12. [PMID: 10207617].
- Ichijo H. From receptors to stress-activated map kinases. Oncogene 1999; 18:6087-93. [PMID: 10557099].
- Roux PP, Blenis J. Erk and p38 mapk-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiology and molecular biology reviews : MMBR 2004; 68:320-44. [PMID: 15187187].
- Peterson RT, Schreiber SL. Translation control: Connecting mitogens and the ribosome. Current biology : CB 1998; 8:R248-50. [PMID: 9545190].
- Jefferies HB, Fumagalli S, Dennis PB, Reinhard C, Pearson RB, Thomas G. Rapamycin suppresses 5'top mrna translation through inhibition of p70s6k. The EMBO journal 1997; 16:3693-704. [PMID: 9218810].
- Iwenofu OH, Lackman RD, Staddon AP, Goodwin DG, Haupt HM, Brooks JS. Phospho-s6 ribosomal protein: A potential new predictive sarcoma marker for targeted mtor therapy. Modern pathology : an official journal of the United States and Canadian Academy of Pathology, Inc 2008; 21:231-7. [PMID: 18157089].
- Hsu SY, Kaipia A, McGee E, Lomeli M, Hsueh AJ. Bok is a pro-apoptotic bcl-2 protein with restricted expression in reproductive tissues and heterodimerizes with selective anti-apoptotic bcl-2 family members. Proc Natl Acad Sci U S A 1997; 94:12401-6. [PMID: 9356461].
- Yang E, Zha J, Jockel J, Boise LH, Thompson CB, Korsmeyer SJ. Bad, a heterodimeric partner for bcl-xl and bcl-2, displaces bax and promotes cell death. Cell 1995; 80:285-91. [PMID: 7834748].
- Inohara N, del Peso L, Koseki T, Chen S, Nunez G. Rick, a novel protein kinase containing a caspase recruitment domain, interacts with clarp and regulates cd95-mediated apoptosis. The Journal of biological chemistry 1998; 273:12296-300. [PMID: 9575181].
- Zha J, Harada H, Yang E, Jockel J, Korsmeyer SJ. Serine phosphorylation of death agonist bad in response to survival factor results in binding to 14-3-3 not bcl-x(l). Cell 1996; 87:619-28. [PMID: 8929531].
- Korsmeyer SJ, Shutter JR, Veis DJ, Merry DE, Oltvai ZN. Bcl-2/bax: A rheostat that regulates an anti-oxidant pathway and cell death. Seminars in cancer biology 1993; 4:327-32. [PMID: 8142617].
- Jones S, Zhang X, Parsons DW, Lin JC, Leary RJ, Angenendt P, Mankoo P, et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2008; 321:1801-6. [PMID: 18772397].
- Hanahan D, Weinberg RA. Hallmarks of cancer: The next generation. Cell 2011; 144:646-74. [PMID: 21376230].
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000; 100:57-70. [PMID: 10647931].










