Keywords
Diabetic embryopathy; Birth defects; Oxidative stress; Endoplasmic reticulum stress; Nitrosative stress; Phytochemical; Polyphenol; Flavonoid
Introduction
Diabetes mellitus in early pregnancy is one of the maternal diseases that cause birth defects in infants, a diabetic complication known as diabetic embryopathy [1,2]. About 10% of newborn babies to diabetic mothers have at least one structural defect, much higher than the background birth defect rate (3%) [3,4]. With a rapid growth of diabetic population, the number of newborn babies having structural anomalies is projected to increase [5]. Therefore, it is urgent to develop therapeutic interventions to prevent birth defects in diabetic pregnancies.
Literature Review
Developmental malformations in the embryo of diabetic pregnancy
Clinical data show that structural defects in infants of diabetic mothers are seen in many organs, and most common in the central nervous and cardiovascular systems (CNS and CVS) [2,6,7]. In the CNS, anencephaly, excencephaly, and microcephaly in the brain and spinal bifida in the spinal cord are frequently seen in infants [2,7]. In the CVS, hypoplastic ventricles, ventricular septal defects, conotruncal anomalies, double outlet ventricles, and tetralogy of Fallot are common in human cases [8,9].
In order to investigate the mechanisms underlying the hyperglycemia-induced fetal abnormalities, animal models of diabetic pregnancy have been generated and used [7,10]. In the commonly used diabetic mouse and rat models, embryonic and fetal structural defects in the CNS and CVS have been observed, similar to those in human infants (Figures 1 and 2) [1,2].
Figure 1: NTDs in the fetuses of diabetic mice. (A) Non-diabetic control; (B) Diabetes, excencephyly; (C) Diabetes, anencephaly. Scale bar=2 mm (apply to all images).
Figure 2: Heart defects in fetuses of diabetic mice. Frontal sections of the heart. (A) Non-diabetic control; (B) Diabetes. The arrow and arrow head indicate septal defects and myocardial hypoplasia, respectively. AO: Aorta, IVS: Interventricular Septum; LV: Left Ventricle; RA: Right Atrium. Scale bar=200 μm.
Glucose as a teratogen in diabetic pregnancy
It has been shown that high glucose is the teratogenic factor, whereas other factors associated with diabetes, such as ketone bodies, advanced glycation end products, and branched chain amino acids, may synergistically affect the action of glucose [11- 13]. Although maintenance of euglycemia in diabetic women before or during the first trimester can reduce the prevalence of fetal abnormalities, to prevent birth defects is still a tough task [14,15]. The reasons for this include that (1) euglycemia is difficult to achieve and maintain even with excellent compliance and clinical care; (2) short episodes of hyperglycemia during critical stages of embryogenesis can cause irreversible damages to the embryo; (3) many women do not know that they are diabetic before conception; and (4) many diabetic women do not plan pregnancies and do not seek prenatal care [16-18]. Therefore, therapeutic interventions are needed to protect embryos from hyperglycemic insult.
Intracellular stresses in diabetic embryopathy
Glucose influx into the embryonic cells disturbs normal intracellular glycolysis, and subsequent protein O-GlcNAcylation, via the as hexosamine biosynthetic pathway pathway, and perturbation of homeostasis in phospholipid metabolism [19,20]. The abnormal intracellular activities impact gene expression and organelle functions. Maternal hyperglycemia upregulates nitric oxide (NO) synthase 2 (Nos2), also known as inducible Nos (iNos), resulting in elevation of NO levels in the embryo [21,22]. High levels of reactive nitrogen species (RNS) lead to increases in protein nitrosylation and nitration and subsequent nitrosative stress [23,24].
Dysfunction of the rough endoplasmic reticulum (ER) causes defects in protein folding. Unfolded and misfolded proteins cannot be processed, packaged, and transported out of the ER, resulting in excessive accumulation of unfolded or misfolded polypeptides in its lumen, generating ER stress [25,26]. First, the UPR increases the expression of chaperone proteins to resolve the protein folding crisis [27,28]. Second, the UPR attenuates protein translation and eliminates the abnormally folded proteins via degradation. Third, if the crisis persists, the UPR arrests mitosis, and/or trigger apoptosis to eliminate the cells [27,29].
Maternal hyperglycemia also alters morphogenesis and function of mitochondria and elevates levels of reactive oxygen species (ROS) in the embryo [30,31]. In addition, it diminishes endogenous antioxidative buffering, including depletion of antioxidants, such as glutathione, and reduction of the expression and activity of antioxidative enzymes, including superoxide dismutases (SODs) and glutathione peroxidases (GPXes), resulting in oxidative stress [32,33].
Interventional studies in animal models
The hyperglycemia-induced intracellular stress conditions have been targets for interventional studies to reduce embryonic malformations in diabetic animal models [1,7]. During the past decades, efforts have been focused on oxidative stress [1,33]. Folate (vitamin B12), a single carbon donor, is involved in DNA synthesis, DNA methylation, and redox regulation [34,35]. Supplementation of folic acid has long been used in perinatal care to reduce fetal abnormalities and other adverse pregnancy outcomes [34,35]. With respect to diabetic embryopathy, addition of folic acid can reduce NTDs in animal embryos cultured in high glucose [36]. Treatment with folic acid in diabetic pregnant animals has also shown to decrease embryonic malformations [36-38]. However, in humans, nearly 30% of pregnancies complicated with diabetes are resistant to folic acid treatment [39]. The underlying mechanisms are still unknown.
A large number of antioxidants, including vitamin C, vitamin E, lipoic acid, ergothioneine, and N-acetylcysteine, have been tested in diabetic animals and all shown to reduce embryonic malformations [40-49]. Arachidonic acid and myo-inositol have been used to restore the deficiency caused by maternal hyperglycemia and to inhibit ROS-mediated lipid peroxidation [40,50,51]. However, the enthusiasm of using antioxidants to prevent birth defects is dampened by the failure of clinical trials to treat similar diseases, such as preeclampsia and cardiovascular diseases [52-54].
Strategies to target other stress conditions are warranted for exploration. Facilitating protein folding to alleviate ER stress is a candidate approach. A number of chemical chaperones, such as phenylbutyric acid (PBA) and tauroursodeoxycholic acid (TUDCA) have been shown to ameliorate diseases in animal models and humans, such as cystic fibrosis and diabetes [55-58]. PBA has been shown to reduce NTDs in mouse embryos in vitro and rescue endocardial cell migration in heart tissues cultured in high glucose [59,60]. More significantly, it has been demonstrated to reduce NTDs in embryos of diabetic mice in vivo [61].
Alleviation of nitrosative stress can be achieved by inhibiting NOS2 activity. Such approach has been demonstrated to be feasible. Oral treatment of diabetic pregnant mice with an NOS2 inhibitor, L-N6-(1-iminoethyl)-lysine, has shown to decrease embryonic malformation rate [62].
Phytochemicals for potential applications in human pregnancies
Because of the unique characteristics of diabetic embryopathy, interventions should be non-invasive, easy to obtain and use before conception, and safe to the embryo and mother. Thus, oral treatment and dietary supplementation are the practical routes of administration. However, the identification of safe and effective therapeutic agents becomes the major obstacle in birth defect prevention.
Attentions have been drawn to naturally occurring products, phytochemicals, in fruits, vegetables, and even traditional medicines [63]. The most common dietary phytochemicals include flavonoids, stilbenoids, and curcuminoids [64-67]. The common characteristic of these phytochemicals is that they possess phenyl rings in their backbone, thus, also known as polyphenols (Tables 1-3). Multiple hydroxyl groups are attached to the phenyl rings, providing active sites for reaction with other molecules [68,69]. These hydroxyl groups can be replaced with other groups, such as methyl and glycoside, to generate a large variety of derivatives (Tables 1-3). Different derivatives have different physical and chemical characteristics and therapeutic properties.
| Common name |
Molecular structure and another name |
| Quercetin |
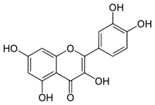
Quercetin aglycone |
| Quercetrin |
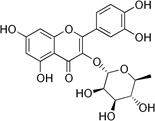
Quercetin-3-L-rhamnoside |
| Rutin |
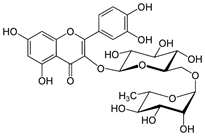
Quercetin-3-O-rutinoside |
| Isoquercetin |
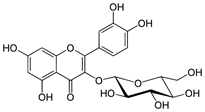
Quercetin-3-O-glucoside |
| Spiraeoside |
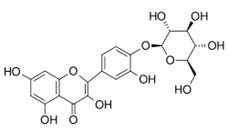
Quercetin-4'-O-glucoside |
Table 1: Major naturally occurring flavonols.
| Common name |
Molecular structure and modification |
| Resveratrol |
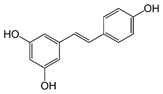
Aglycone |
| Pterosilbene |
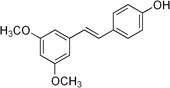
Aglycone |
| Piceatannol |
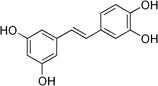
Aglycone |
| Astringin |
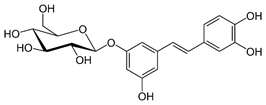
Glycoside |
| Piceid |
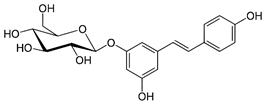
Glycoside |
Table 2: Major naturally occurring stilbenoids.
| Common name |
Molecular structure |
| Curcumin |
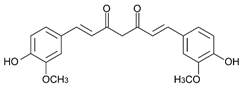 |
| Bisdemethoxycurcumin |
 |
| Demethoxycurcumin |
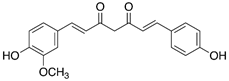 |
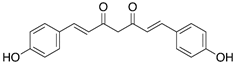
Table 3: Major naturally occurring curcuminoids.
Resveratrol, a stilbenoid antioxidant, has been shown to reduce apoptosis in the embryos of diabetic animals and restore the levels of proteins that are involved in regulation of apoptosis and development, including retinoic acid receptors and protein kinases [70,71]. Curcumin, rich in turmeric, and punicalagin, found in pomegranate juice, can also reduce NTDs in mouse embryos cultured in high glucose [72,73].
Discussion
Derivatives of flavonoid also possess antioxidative properties. Epigallocatechin-3-gallate (EGCG), abundant in green tea, has been shown to reduce NTDs in mouse embryos in vitro and in vivo [74,75]. In addition to antioxidative properties, some of polyphenols also inhibit NOS expression and activity and block ER stress-activated molecular pathways [76-78]. Among the flovonols, quercetin (QC) has been shown to reduce NTDs in the embryos of diabetic mice. Such effect is associated with alleviation of nitrosative stress, indicated by decreases in protein nitrosylation and nitration, and oxidative stress with reduced levels of lipoperoxation products [79]. QC upregulates genes that encode antioxidative enzymes to enhance endogenous antioxidative capacity for long-lasting protection against oxygen free radicals [79]. Moreover, QC also increases the expression of the genes involved in DNA damage repair [79]. These suggest that QC affects multiple molecular systems to enforce cellular defense against hyperglycemic insult.
However, QC is an aglycone form of flavonoid. It has lower solubility in water and lower rate of absorption in the gastrointestinal tract (GI) than its modified derivatives [80,81]. These characteristics may limit its applicability in clinical practice.
Quercetin-3-glycoside (Q3G), also known as isoquercetin, is a glucoside derivative of QC. It has higher water solubility and higher GI absorption rate [80,82,83]. Q3G is relatively stable in circulation and thus, has higher bioavailability in organ systems. Unlike other QC derivatives, Q3G can be transported via glucose carriers, which increases its bioavailibility to cells [84]. Treatment with Q3G in vivo reduces NTDs in the embryos of diabetic mice [85]. While alleviating intracellular stress conditions, Q3G modulates the signaling of the NFκB system, which is involved in regulating the expression of NOSes and SODs [85].
Conclusion
Human pregnancy presents unique challenges for intervention to prevent birth defects. The safety and effectiveness are the key factors in development of intervention strategies. Searching for safe and effective agents from phytochemicals is an approach of worth taking. Vagarous studies are required to delineate the pharmacodynamics and pharmacokinetics of candidate agents in both maternal and embryonic systems. Complete understanding of the cellular and molecular mechanisms underlying their actions in diabetic embryopathy is also essential for ensure the feasibility in application in human pregnancies.
References
- Zhao Z, Reece EA (2013) New concepts in diabetic embryopathy. Clin Lab Med 33: 207-233.
- Correa A, Gilboa SM, Besser LM, Botto LD, Moore CA, et al. (2008) Diabetes mellitus and birth defects. Am J Obstet Gynecol 199: e231-239.
- Centers for Disease Control and Prevention (2008) Update on overall prevalence of major birth defects-Atlanta, Georgia, 1978-2005. Morb Mort Wkly Rep 1: 57.
- Yoon PW, Rasmussen SA, Lynberg MC, Moore CA, Anderka M, et al. (2001) The national birth defects prevention study. Public Health Rep 116: 32-40.
- http: //wwwcdcgov/diabetes/pubs/statsreport14/national-diabetes-report-webpdf.
- Reece EA, Eriksson UJ (1996) The pathogenesis of diabetes-associated congenital malformations. Obstet Gynecol Clin North Am 23: 29-45.
- Zhao Z, Reece EA (2005) Experimental mechanisms of diabetic embryopathy and strategies for developing therapeutic interventions. J Soc Gynecol Investig 12: 549-557.
- Aberg A, Westbom L, Kallen B (2001) Congenital malformations among infants whose mothers had gestational diabetes or preexisting diabetes. Early Hum Dev 61: 85-95.
- Corrigan N, Brazil DP, McAuliffe F (2009) Fetal cardiac effects of maternal hyperglycemia during pregnancy. Birth Defects Res Clin Mol Teratol 85: 523-530.
- Yang P, Zhao Z, Reece EA (2008) Activation of oxidative stress signaling that is implicated in apoptosis with a mouse model of diabetic embryopathy. Am J Obstet Gynecol 198: 130 e131-137.
- Greene MF, Hare JW, Cloherty JP, Benacerraf BR, Soeldner JS (1989) First-trimester hemoglobin A1 and risk for major malformation and spontaneous abortion in diabetic pregnancy. Teratology 39: 225-231.
- Miller E, Hare JW, Cloherty JP, Dunn PJ, Gleason RE, et al. (1981) Elevated maternal hemoglobin A1c in early pregnancy and major congenital anomalies in infants of diabetic mothers. N Engl J Med 304: 1331-1334.
- Eriksson UJ, Borg LA, Cederberg J, Nordstrand H, Siman CM, et al. (2000) Pathogenesis of diabetes-induced congenital malformations. Ups J Med Sci 105: 53-84.
- Fuhrmann K, Reiher H, Semmler K, Glockner E (1984) The effect of intensified conventional insulin therapy before and during pregnancy on the malformation rate in offspring of diabetic mothers. Exp Clin Endocrinol 83: 173-177.
- Kitzmiller JL, Gavin LA, Gin GD, Jovanovic-Peterson L, Main EK, et al. (1991) Preconception care of diabetes: Glycemic control prevents congenital anomalies. JAMA 265: 731-736.
- Mills JL, Baker L, Goldman AS (1979) Malformations in infants of diabetic mothers occur before the seventh gestational week: Implications for treatment. Diabetes 28: 292-293.
- Holing EV, Beyer CS, Brown ZA, Connell FA (1998) Why don't women with diabetes plan their pregnancies? Diabetes Care 21: 889-895.
- Lipscombe LL, McLaughlin HM, Wu W, Feig DS (2011) Pregnancy planning in women with pregestational diabetes. J Matern Fetal Neonatal Med 24: 1095-1101.
- Kim G, Cao L, Reece EA, Zhao Z (2017) Impact of protein O-GlcNAcylation on neural tube malformation in diabetic embryopathy. Sci Rep 7: 11107.
- Cao L, Liu P, Gill K, Reece EA, Cheema AK, et al. (2016) Identification of novel cell survival regulation in diabetic embryopathy via phospholipidomic profiling. Biochem Biophys Res Commun 470: 599-605.
- Sugimura Y, Murase T, Oyama K, Uchida A, Sato N, et al. (2009) Prevention of neural tube defects by loss of function of inducible nitric oxide synthase in fetuses of a mouse model of streptozotocin-induced diabetes. Diabetologia 52: 962-971.
- White V, Gonzalez E, Pustovrh C, Capobianco E, Martinez N, et al. (2007) Leptin in embryos from control and diabetic rats during organogenesis: A modulator of nitric oxide production and lipid homeostasis. Diabetes Metab Res Rev 23: 580-588.
- Hess DT, Matsumoto A, Kim SO, Marshall HE, Stamler JS (2005) Protein S-nitrosylation: Purview and parameters. Nat Rev Mol Cell Biol 6: 150-166.
- Knott AB, Bossy-Wetzel E (2009) Nitric oxide in health and disease of the nervous system. Antioxid Redox Signal 11: 541-554.
- Eizirik DL, Cardozo AK, Cnop M (2008) The role for endoplasmic reticulum stress in diabetes mellitus. Endocr Rev 29: 42-61.
- Malhotra JD, Kaufman RJ (2007) The endoplasmic reticulum and the unfolded protein response. Semin Cell Dev Biol 18: 716-731.
- Hegde RS, Ploegh HL (2010) Quality and quantity control at the endoplasmic reticulum. Curr Opin Cell Biol 22: 437-446.
- Scheper W, Hoozemans JJ (2009) Endoplasmic reticulum protein quality control in neurodegenerative disease: the good, the bad and the therapy. Curr Med Chem 16: 615-626.
- Marciniak SJ, Ron D (2006) Endoplasmic reticulum stress signaling in disease. Physiol Rev 86: 1133-1149.
- Yang X, Borg LA, Eriksson UJ (1995) Altered mitochondrial morphology of rat embryos in diabetic pregnancy. Anat Rec 241: 255-267.
- Yang X, Borg LA, Eriksson UJ (1997) Altered metabolism and superoxide generation in neural tissue of rat embryos exposed to high glucose. Am J Physiol 272: E173-E180.
- Loeken MR (2004) Free radicals and birth defects. J Matern Fetal Neonatal Med 15: 6-14.
- Yang P, Reece EA, Wang F, Gabbay-Benziv R (2015) Decoding the oxidative stress hypothesis in diabetic embryopathy through proapoptotic kinase signaling. Am J Obstet Gynecol 212: 569-579.
- Beaudin AE, Stover PJ (2007) Folate-mediated one-carbon metabolism and neural tube defects: Balancing genome synthesis and gene expression. Birth Defects Res C Embryo Today 81: 183-203.
- Butterworth CE, Bendich A (1996) Folic acid and the prevention of birth defects. Annu Rev Nutr 16: 73-97.
- Wentzel P, Gareskog M, Eriksson UJ (2005) Folic acid supplementation diminishes diabetes- and glucose-induced dysmorphogenesis in rat embryos in vivo and in vitro. Diabetes 54: 546-553.
- Oyama K, Sugimura Y, Murase T, Uchida A, Hayasaka S, et al. (2009) Folic acid prevents congenital malformations in the offspring of diabetic mice. Endocr J 56: 29-37.
- Yuan Q, Zhao S, Liu S, Zhang Y, Fu J, et al. (2013) Folic acid supplementation changes the fate of neural progenitors in mouse embryos of hyperglycemic and diabetic pregnancy. J Nutr Biochem 24: 1202-1212.
- Mersereau PK, Carter K, Fassett H, Williams E, Flores GJ, et al. (2004) Spina bifida and anencephaly before and after folic acid mandate-United States, 1995-1996 and 1999-2000. MMWR Morb Mortal Wkly Rep 53: 362-365.
- Reece EA, Wu YK, Zhao Z, Dhanasekaran D (2006) Dietary vitamin and lipid therapy rescues aberrant signaling and apoptosis and prevents hyperglycemia-induced diabetic embryopathy in rats. Am J Obstet Gynecol 194: 580-585.
- Siman CM, Eriksson UJ (1997) Vitamin E decreases the occurrence of malformations in the offspring of diabetic rats. Diabetes 46: 1054-1061.
- Siman CM, Eriksson UJ (1997) Vitamin C supplementation of the maternal diet reduces the rate of malformation in the offspring of diabetic rats. Diabetologia 40: 1416-1424.
- Sivan E, Reece EA, Wu YK, Homko CJ, Polansky M, et al. (1996) Dietary vitamin E prophylaxis and diabetic embryopathy: Morphologic and biochemical analysis. Am J Obstet Gynecol 175: 793-799.
- Viana M, Herrera E, Bonet B (1996) Teratogenic effects of diabetes mellitus in the rat: Prevention by vitamin E. Diabetologia 39: 1041-1046.
- Zaken V, Kohen R, Ornoy A (2001) Vitamins C and E improve rat embryonic antioxidant defense mechanism in diabetic culture medium. Teratology 64: 33-44.
- Wiznitzer A, Ayalon N, Hershkovitz R, Khamaisi M, Reece EA, et al. (1999) Lipoic acid prevention of neural tube defects in offspring of rats with streptozocin-induced diabetes. Am J Obstet Gynecol 180: 188-193.
- Al-Ghafli MH, Padmanabhan R, Kataya HH, Berg B (2004) Effects of alpha-lipoic acid supplementation on maternal diabetes-induced growth retardation and congenital anomalies in rat fetuses. Mol Cell Biochem 261: 123-135.
- Moazzen H, Lu X, Ma NL, Velenosi TJ, Urquhart BL, et al. (2014) N-Acetylcysteine prevents congenital heart defects induced by pregestational diabetes. Cardiovasc Diabetol 13: 1-13.
- Wentzel P, Eriksson UJ (1998) Antioxidants diminish developmental damage induced by high glucose and cyclooxygenase inhibitors in rat embryos in vitro. Diabetes 47: 677-684.
- Reece EA, Wu YK (1997) Prevention of diabetic embryopathy in offspring of diabetic rats with use of a cocktail of deficient substrates and an antioxidant. Am J Obstet Gynecol 176: 790-797.
- Khandelwal M, Reece EA, Wu YK, Borenstein M (1998) Dietary myo-inositol therapy in hyperglycemia-induced embryopathy. Teratology 57: 79-84.
- Villar J, Purwar M, Merialdi M, Zavaleta N, Ngoc N, et al. (2009) World Health Organisation multicentre randomised trial of supplementation with vitamins C and E among pregnant women at high risk for pre-eclampsia in populations of low nutritional status from developing countries. Br J Obstet Gynecol 116: 780-788.
- Polyzos NP, Mauri D, Tsappi M, Tzioras S, Kamposioras K, et al. (2007) Combined vitamin C and E supplementation during pregnancy for preeclampsia prevention: a systematic review. Obstet Gynecol Surv 62: 202-206.
- Briasoulis A, Tousoulis D, Antoniades C, Stefanadis C (2009) The oxidative stress menace to coronary vasculature: any place for antioxidants?. Curr Pharm Des 15: 3078-3090.
- Kars M, Yang L, Gregor MF, Mohammed BS, Pietka TA, et al. (2010) Tauroursodeoxycholic Acid may improve liver and muscle but not adipose tissue insulin sensitivity in obese men and women. Diabetes 59: 1899-1905.
- Roomans GM (2001) Pharmacological treatment of the ion transport defect in cystic fibrosis. Expert Opin Investig Drugs 10: 1-19.
- Engin F, Yermalovich A, Nguyen T, Hummasti S, Fu W, et al. (2013) Restoration of the unfolded protein response in pancreatic beta cells protects mice against type 1 diabetes. Sci Transl Med 5: 211ra156.
- Ozcan U, Yilmaz E, Ozcan L, Furuhashi M, Vaillancourt E, et al. (2006) Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science 313: 1137-1140.
- Li X, Xu C, Yang P (2013) c-Jun NH2-terminal kinase 1/2 and endoplasmic reticulum stress as interdependent and reciprocal causation in diabetic embryopathy. Diabetes 62: 599-608.
- Zhao Z (2012) Endoplasmic reticulum stress in maternal diabetes-induced cardiac malformations during critical cardiogenesis period. Birth Defects Res B Dev Reprod Toxicol 95: 1-6.
- Zhao Z, Cao L, Reece EA (2017) Formation of neurodegenerative aggresome and death-inducing signaling complex in maternal diabetes-induced neural tube defects. Proc Natl Acad Sci USA 114: 4489-4494.
- Zhao Z, Eckert RL, Reece EA (2012) Reduction in embryonic malformations and alleviation of endoplasmic reticulum stress by nitric oxide synthase inhibition in diabetic embryopathy. Reprod Sci 19: 823-831.
- Krzyzanowska J, Czubacka A, Oleszek W (2010) Dietary phytochemicals and human health. Adv Exp Med Biol 698: 74-98.
- Xiao ZP, Peng ZY, Peng MJ, Yan WB, Ouyang YZ, et al. (2011) Flavonoids health benefits and their molecular mechanism. Mini Rev Med Chem 11: 169-177.
- Das S, Das DK (2007) Resveratrol: A therapeutic promise for cardiovascular diseases. Recent Pat Cardiovasc Drug Discov 2: 133-138.
- Singh BN, Shankar S, Srivastava RK (2011) Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem Pharmacol 82: 1807-1821.
- Gupta SC, Patchva S, Koh W, Aggarwal BB (2012) Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clinical and experimental pharmacology & physiology 39: 283-299.
- Veitch NC, Grayer RJ (2008) Flavonoids and their glycosides, including anthocyanins. Nat Prod Rep 25: 555-611.
- Cazarolli LH, Zanatta L, Alberton EH, Figueiredo MS, Folador P, et al. (2008) Flavonoids: Prospective drug candidates. Mini Rev Med Chem 8: 1429-1440.
- Singh CK, Kumar A, Hitchcock DB, Fan D, Goodwin R, et al. (2011) Resveratrol prevents embryonic oxidative stress and apoptosis associated with diabetic embryopathy and improves glucose and lipid profile of diabetic dam. Mol Nutr Food Res 55: 1186-1196.
- Singh CK, Kumar A, LaVoie HA, DiPette DJ, Singh US (2012) Resveratrol prevents impairment in activation of retinoic acid receptors and MAP kinases in the embryos of a rodent model of diabetic embryopathy. Reprod Sci 19: 949-961.
- Wu Y, Wang F, Reece EA, Yang P (2015) Curcumin ameliorates high glucose-induced neural tube defects by suppressing cellular stress and apoptosis. Am J Obstet Gynecol
- Zhong J, Reece EA, Yang P (2015) Punicalagin exerts protective effect against high glucose-induced cellular stress and neural tube defects. Biochem Biophys Res Commun 467: 179-184.
- Yang P, Li H (2010) Epigallocatechin-3-gallate ameliorates hyperglycemia-induced embryonic vasculopathy and malformation by inhibition of Foxo3a activation. Am J Obstet Gynecol 203: 75 e71-76.
- Zhong J, Xu C, Reece EA, Yang P (2016) The green tea polyphenol EGCG alleviates maternal diabetes-induced neural tube defects by inhibiting DNA hypermethylation. Am J Obstet Gynecol 215: 368 e361-368 e310.
- Mladenka P, Zatloukalova L, Filipsky T, Hrdina R (2010) Cardiovascular effects of flavonoids are not caused only by direct antioxidant activity. Free Radic Biol Med 49: 963-975.
- Bakhshi J, Weinstein L, Poksay K, Nishinaga B, Bredesen D, et al. (2008) Coupling endoplasmic reticulum stress to the cell death program in mouse melanoma cells: Effect of curcumin. Apoptosis 13: 904-914.
- Chinta SJ, Poksay KS, Kaundinya G, Hart M, Bredesen DE, et al. (2009) Endoplasmic reticulum stress-induced cell death in dopaminergic cells: Effect of resveratrol. J Mol Neurosci 39: 157-168.
- Cao L, Tan C, Meng F, Liu P, Reece EA, et al. (2016) Amelioration of intracellular stress and reduction of neural tube defects in embryos of diabetic mice by phytochemical quercetin. Sci Rep 6: 21491.
- Morand C, Manach C, Crespy V, Remesy C (2000) Respective bioavailability of quercetin aglycone and its glycosides in a rat model. Biofactors 12: 169-174.
- Kelly GS (2011) Quercetin monograph. Altern Med Rev 16: 172-194.
- Hollman PC, Katan MB (1999) Health effects and bioavailability of dietary flavonols. Free Radic Res 31: S75-80.
- Day AJ, Gee JM, DuPont MS, Johnson IT, Williamson G (2003) Absorption of quercetin-3-glucoside and quercetin-4'-glucoside in the rat small intestine: The role of lactase phlorizin hydrolase and the sodium-dependent glucose transporter. Biochem Pharmacol 65: 1199-1206.
- Gee JM, DuPont MS, Rhodes MJ, Johnson IT (1998) Quercetin glucosides interact with the intestinal glucose transport pathway. Free Radic Biol Med 25: 19-25.
- Tan C, Meng F, Reece EA, Zhao Z (2018) Modulation of nuclear factor-kappaB signaling and reduction of neural tube defects by quercetin-3-glucoside in embryos of diabetic mice. Am J Obstet Gynecol 2: 1.