Keywords
Two-component signal-transduction systems; Histidine kinase; Response regulator; Antibacterials
Introduction
Bacteria are wildly spread in nature, and some of them are human pathogen. To survive or even persistently infect in several intricate host environments, bacteria need adapt to the environment. Organisms are able to sense environmental signals and transduce the signals to the intracellular to initiate the process of transcription, translation, protein expression, modification and other biological processes of related genes. The process of sensing the environment and making adaptive response to external stimuli is often mediated by two-component signal-transduction systems (TCSTSs). The signal transduction system is activated by external stimuli including temperature, nutrients and pH [1].
Two component systems are very common in nature. This system has been considered to be restricted in prokaryotes for a long time. However, the emerging evidences indicate that this system also exists in many eukaryotes, including yeast, fungi, slime molds, and higher plants. But the system so far has not been found in vertebrates. It is firstly identified by Ninfa and Magasanik when they studied nitrogen regulatory protein of Escherichia coli ( E. coil) in 1986 [2]. Meanwhile, in 1986 Nixon found that there were many sensory systems in bacteria. By comparing the amino acid sequences, they found that these sensory system components were similar to the nitrogen regulatory system of E. coil [3].
Representative TCSTS consists of two different types of protein, a histidine kinase (HK) and a response regulator (RR). The HK is responsible for responding to external stimuli by autophosphorylation, subsequently transfers this phosphate group to the RR. This phosphorylation changes the characters of RR protein and enables the RR to bind to the promoter region of target gene(s), functioning as a transcriptional factor [4] (Figure 1). These signal transduction pathways need phosphorylase to dephosphorylate and restore the unstimulated state of RR, thus the system can respond to the external stimuli again. The phosphorylase can be a histidine kinase itself, a response regulator, or one of the other proteins.
Based on genomic sequencing, a lot of gene families have been identified. The two component system gene is firstly demonstrated by this method. Researchers have made extensive studies on various two component systems, particularly which plays an important role in cell physiology, information transfer, and individual growth in bacterial pathogens. Moreover two component systems have been shown in a number of cases to regulate virulence factors, which are crucial for the pathogenicity of many bacterial species [5], competence [6], biofilm formation [7-9] and cell envelope response [10], and antibiotics resistance [5] (Figure 2). Indeed, reports have shown that inhibitors of two component systems have good antibacterial activity [11]. Two component systems have four similar characters which may make the system to be a good potential target for antimicrobials. The characters are as follows: ? The HK and RR proteins have significant homology respectively in different genera of bacteria, especially in the residues residing near active sites [12]; ? The two-component signal transduction system can regulate the expression of the pathogen essential virulence factors that are required for survival in the host use to regulate of essential virulence factors [13-15]; ? There are many two component systems in bacteria, and some are vital for viability [16,17]; ? The signal transduction in mammals has an entirely different mechanism. Taken together, this family of proteins may be very suited as a target for the development of novel antibacterials.
TCSTSs Protein Structure
The HK is located in the cell membrane and usually has one or more transmembrane regions. The HK forms a dimer to fulfill functions and its C-terminal contains a hist???idine kinase domain (about 200 residues). Autophosphorylation process happens in this domain and the phosphorylation sites are generally conserved histidine residues. The sequence alignment showed that this domain has five unique conserved regions: H, N, D, F and G [12]. One module that can sense the external signal is named as input module and is connected with histidine kinase domain. The N-terminal of HK is very different, usually comprising several transmembrane domains. A linker connects the histidine kinase domain and the transmembrane domain (Figure 2B). These structure features make the HK can sensitively sense the change of the external environment.
The RR protein resides in the cytoplasm, and has a highly conserved receiver domain in its N-terminal region (about 110 amino acids) by comparing different RR proteins and an effector or output domain within the C-terminal part that can mostly bind DNA (Figure 2C). The phosphorylation sites are aspartic acid residues. Some of the RR proteins are transcription regulators. The signal transduction process is composed of the signal input, histidine autophosphorylation, response regulator protein phosphorylation and signal output component.
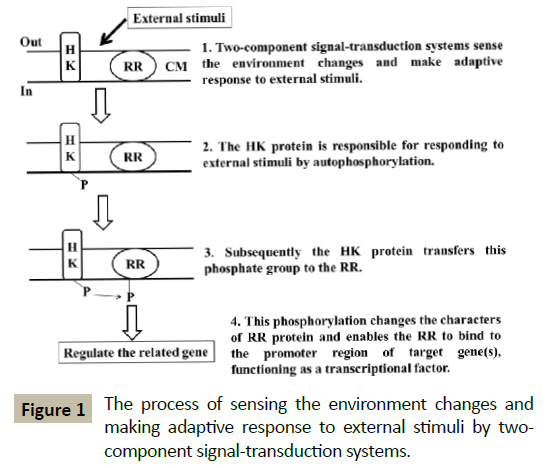
Figure 1: The process of sensing the environment changes and making adaptive response to external stimuli by twocomponent signal-transduction systems.
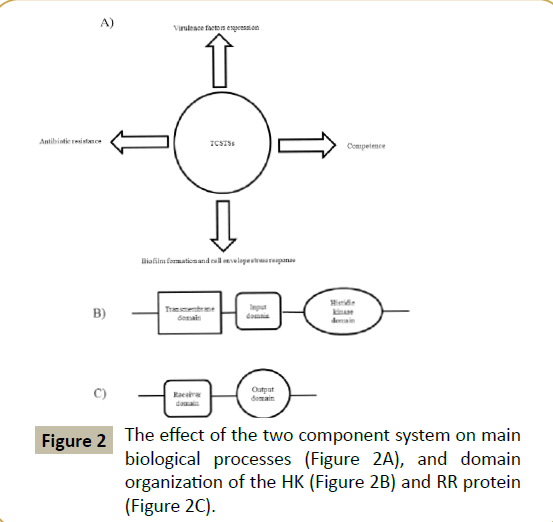
Figure 2:The effect of the two component system on main biological processes (Figure 2A), and domain organization of the HK (Figure 2B) and RR protein (Figure 2C).
Regulation of Virulence Factors Expression
Virulence is very crucial in the infection and pathogenesis of the pathogens. The regulation of virulence factors expression is controlled by many network systems and the two component system is one of them. Based on genomic sequencing, many putative HK proteins, with one ‘orphan’ RR proteins have been found in bacteria. For example, 31, 13 and 14 two component systems are found in E. coli, Streptococcus pneumoniae (S. pneumoniae) and Streptococcus mutans (S. mutans) respectively [18-21]. Some of them have been proved that they can regulate the expression of virulence factors. LiaS, one of two component system proteins in S. mutans, negatively regulates of the expression of GbpC, which is a very important surface-associated protein. LiaS binds dextran and facilitates the biofilm formation of the organism, and adherence and accumulation on the tooth. In addition, LiaS activates the production of mutacin IV, which is under the control of a common regulatory network along with gbpC [22]. The two component family member HK/RR04 has been reported to relate to the regulation of the manganese transporter PsaA [23]. Moreover, another two component system protein HK/RR06 regulates the expression of the virulence factor CbpA [24]. The CiaRH system has been shown that it influences the expression of high-temperature requirement A (HtrA), which is a serine protease and plays vital roles in protein quality control in prokaryotes and eukaryotes [25,26]. The ArlRS twocomponent system of Staphylococcus aureus is related to the expression of virulence factors such as cell-wall-associated protein A. The deletion of ArlRS attenuates the effect of DNA supercoiling regulators on protein A expression, indicating that ArlRS system indirectly regulates the expression of protein A [27]. BlpRH two component systems is linked to the production of barteriocin in S. pneumoniae [4]. Two component system protein RitR negatively regulates the piu hemin-iron transporter system of S. pneumoniae, thus suppresses the uptake of iron [28]. The two component system protein RR09 contributes to the virulence of S. pneumoniae in the manner of serotype-specific [29]. The two component system VicR/K may mediate the expression of the virulence factor PspA of S. pneumoniae [30]. These findings provide conclusive evidence for regulation of the virulence of the two component systems and their contribution to the virulence of the organism.
Competence
The ability of naturally acquiring exogenous DNA is known as “competence for transformation” among more than 70 bacterial species [31]. Competence has been extensively shown to form transiently in lag phase [32] or during exponential growth [33-37]. And competence is involved in DNA transport, recombination [38] and repair [39], and/or nutritional requirements via supplying a carbon, nitrogen, phosphorus, and energy source [40]. Competence for genetic transformation of bacteria is reported to be regulated by the two component system ComD/E [6-9], which activates the early competence genes in response to a competence-stimulating peptide [12,13]. The precursor of the competence-stimulating peptide is encoded by the comC gene [6,41,42], and secreted to the extracellular after modification, acting on the ComDE system to trigger expression of related genes [43]. Deletion of comE eliminates both response to synthetic activator peptide and endogenous competence induction [6]. ComE (R120S) overexpression activates ComD-dependent transformation in the absence of the competence-stimulating peptide in S. pneumoniae [44]. There is a correlation between the level of comCDE transcripts and developmental transformability in culture. In addition, another two component system CiaRH is linked to competence regulation [5]. The mutation of CiaH results in competence deficiency of S. pneumoniae, suggesting that CiaH modulates the competence development [45]. And two component systems HK/RR11, has been reported to be related to genetic competence in S. mutans [7-9]. Moreover, the two component system VicRK partly regulates the competence-stimulating peptide (CSP) production in S. mutans [46]. Furthermore, the two component system protein Chis controls a novel transmembrane regulator TfoS by post-translational regulation, whose inactivation abolishes the competence [47]. PnpR/S, a PhoP/Q-like two component, also regulates competence of S. pneumoniae [19,25]. These results indicate that the two component system modulates competence of bacteria to transport and recombine DNA.
Effect on Biofilm Formation and Cell Envelope Stress Response
Two component systems, named ComDE and HK/RR11, have been shown to be involved in biofilm formation. Loss-of-function mutation of hk11 or rr11 leads to defects in biofilm formation, which presents reduced biomass, sponge-like architecture with large gaps, and longer chains composition of cells than those of the parent biofilm in S. mutans [7-9]. Maintaining integrity of cell envelope is crucial for bacterial survival. As reported, the two component system LiaRS of Bacillus subtilis is a part of the regulatory network of cell wall stress response and is critical for maintaining integrity of the bacterial cell wall [48]. LiaR can regulate cell wall synthesis, pili formation and cell membrane modification in group B Streptococcus (GBS) [49]. And the VicRK system modulates several genes associated with cell membrane and cell wall homeostasis in S. pneumoniae [10]. The liaFSR genes mutations lead to the susceptibility to Lipid II cycle interfering antibiotics and chemicals that influenced the cell membrane integrity. Significantly, the LiaFSR system is reported to activate transcription of several genes participating in membrane protein synthesis, peptidoglycan biosynthesis, envelope chaperone/ proteases, and transcriptional regulators under bacitracin stress conditions. These findings demonstrate that LiaFSR system can sense cell envelope stress and preserve envelope integrity in S. mutans [50]. These data demonstrate that two component systems participate in biofilm formation and cell envelope response of bacteria.
Effect on Antibiotic Resistance
The deletion of LiaR is more sensitive to cell wall-active antibiotics (vancomycin and bacitracin) and antimicrobial peptides (polymixin B, colistin, and nisin) compared to wild-type GBS. And mouse models of both GBS sepsis and pneumoniae with LiaR mutant GBS significantly attenuates, suggesting that LiaR controls expression of genes associated with microbial defense against host antimicrobial systems [49]. A LiaR deletion mutant results in reversion of daptomycin and telavancin resistance, as well as leads to hypersensit???ive to these antibiotics and several antimicrobial peptides, indicating that LiaR is a major regulator to control the cell membrane in response to different antibacterial agent and polypeptide in daptomycin-resistant Enterococci faecalis [51]. A T/A deletion mutation of CiaH, resulting in a truncation protein, reverse the resistance to cefotaxime in S. pneumoniae [5,45]. And CiaRH mutants have high defense against the effect of cell wall inhibitors, including cycloserine, bacitracin, and vancomycin, as well as less sensitive to these drugs [52]. LiaRS of Listeria monocytogenes (L. monocytogenes) can be activated by the antibiotics which act on the cell wall, LiaR phosphorylation level is controlled by the LiaS group of protein kinase and phosphatase dual activity. Phosphatase activity of LiaS in the absence of cell wall stress is activated by LiaF [53]. The transcription of LiaR gene is increased in L. monocytogenes under salt stress. And LiaR mutation strains are markedly more susceptible to nisin in comparison with the salt-induced nisin resistance of wild-type strains, suggesting that LiaFSR results in cross-protection and resistance to nisin [54]. These reports manifest that the two component system plays an important role in antibiotic resistance of bacteria.
Discussion
The results discussed above suggest that the two component signal system is very vital to bacteria, involving in the regulation of virulence factor expression, the control of competence, modulation of biofilm formation and cell envelope stress response, and antibiotic resistance. Bacteria are wildly found in nature and two component systems are widespread in bacteria. There are significant homologies between HK, RR proteins and different genera of bacteria respectively, especially in the residues located near active sites. Furthermore the signal transduction in mammals has an entirely different mechanism compared with bacteria. Moreover inhibitors of two component systems have been proved that they have good antimicrobial activity. Altogether, this system is potentially very suited as a good drug target for the development of novel antibacterials. As such, studies on the regulation mechanism of this system on virulence, competence and antibiotic resistance have great significance.
Acknowledgement
This work was supported by the Anhui Provincial Natural Science Foundation (Grant No. 1508085QC49), the school fund project of Anhui Medical University (Grant No. 2015XKJ031) and the doctoral research fund project of the Second Affiliated Hospital of Anhui Medical University (Grant No.2014BKJ034).
References
- Stock BJ, Surette MG, Levit M, Park P (1995) Two-component signal transduction systems: structure?function relationship and mechanisms of catalysis. In: Two-Component Signal Transduction. American Society for Microbiology, DC, USA.
- Ninfa AJ, Magasanik B (1986) Covalent modification of the glnG product, NRI, by the glnL product, NRII, regulates the transcription of the glnALG operon in Escherichia coli. Proc Natl Acad Sci U S A 83: 5909-5913.
- Nixon BT, Ronson CW, Ausubel FM (1986) Two-component regulatory systems responsive to environmental stimuli share strongly conserved domains with the nitrogen assimilation regulatory genes ntrB and ntrC. Proc Natl Acad Sci USA 83: 7850-7854.
- de Saizieu A, Gardes C, Flint N, Wagner C, Kamber M, et al. (2000) Microarray-based identification of a novel Streptococcus pneumoniaeregulon controlled by an autoinduced peptide. J Bacteriol 182: 4696-4703.
- 5. Guenzi E, Gasc AM, Sicard MA, Hakenbeck R (1994) A two component signal-transducing system is involved in competence and penicillin susceptibility in laboratory mutants of Streptococcus pneumoniae. MolMicrobiol 12: 505-515.
- Pestova EV, Havarstein LS, Morrison DA (1996) Regulation of competence for genetic transformation in Streptococcus pneumoniae by an autoinduced peptide pheromone and a two- component regulatory system. MolMicrobiol 21: 853-862.
- Li YH, Hanna MN, Svens?ter G, Ellen RP, Cvitkovitch DG (2001) Cell density modulates acid adaptation in Streptococcus mutans: implications for survival in biofilms. J Bacteriol 183: 6875-6884.
- Li YH, Lau PC, Lee JH, Ellen RP, Cvitkovitch DG (2001) Natural genetic transformation of Streptococcus mutans growing in biofilms. J Bacteriol 183: 897-908.
- Li YH, Lau PC, Tang N, Svens?ter G, Ellen RP, et al. (2002) Novel two-component regulatory system involved in biofilm formation and acid resistance in Streptococcus mutans. J Bacteriol 184: 6333-6342.
- Ng WL, Tsui HC, Winkler ME (2005) Regulation of the pspA virulence factor and essential pcsBmurein biosynthetic genes by the phosphorylated VicR (YycF) response regulator in Streptococcus pneumoniae. J Bacteriol1871: 7444-7459.
- Barrett JF, Goldschmidt RM, Lawrence LE, Foleno B, Chen R, et al. (1998) Antibacterial agents that inhibit two-component signal transduction systems. Proc Natl Acad Sci U S A 95: 5317-5322.
- Parkinson JS, Kofoid EC (1992) Communication modules in bacterial signaling proteins. Annu Rev Genet 26: 71-112.
- Dziejman M, Mekalanos JJ, Hoch JA, Silhavy TJ (1995) Two-component signal transduction and its role in the expression of bacterial virulence factors. In: Hoch JA, Silhavy TJ (edr)Two-Component Signal Transduction. American Society for Microbiology, DC, USA.
- Groisman EA, Heffron F, Hoch JA, Silhavy TJ (1995) In: Hoch JA, Silhavy TJ (edr) Two-Component Signal Transduction. American Society for Microbiology, DC, USA.
- Uh MA, Miller JF, Hoch JA, Silhavy TJ (1995) In: Hoch JA , Silhavy TJ (edr)Two-Component Signal Transduction, American Society for Microbiology, DC, USA.
- Hecht GB, Lane T, Ohta N, Sommer JM, Newton A (1995) An essential single domain response regulator required for normal cell division and differentiation in Caulobactercrescentus. EMBO J 14: 3915-3924.
- Quon KC, Marczynski GT, Shapiro L (1996) Cell cycle control by an essential bacterial two-component signal transduction protein. Cell 84: 83-93.
- Lange R, Wagner C, de Saizieu A, Flint N, Molnos J, et al. (1999) Domain organization and molecular characterization of 13 two-component systems identified by genome sequencing of Streptococcus pneumoniae. Gene237: 223-234.
- Throup JP, Koretke KK, Bryant AP, Ingraham KA, Chalker AF, et al. (2000) A genomic analysis of two-component signal transduction in Streptococcus pneumoniae. MolMicrobiol 35: 566-576.
- Ajdi?? D, McShan WM, McLaughlin RE, Savi?? G, Chang J, et al. (2002) Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc Natl Acad Sci U S A 99: 14434-14439.
- Biswas I, Drake L, Erkina D, Biswas S (2008) Involvement of sensor kinases in the stress tolerance response of Streptococcus mutans. J Bacteriol 190: 68-77.
- Chong P, Drake L, Biswas I (2008) LiaS regulates virulence factor expression in Streptococcus mutans. Infect Immun 76: 3093-3099.
- McCluskey J, Hinds J, Husain S, Witney A, Mitchell TJ (2004) A two-component system that controls the expression of pneumococcal surface antigen A (PsaA) and regulates virulence and resistance to oxidative stress in Streptococcus pneumoniae. MolMicrobiol 51: 1661-1675.
- Standish AJ, Stroeher UH, Paton JC (2005) The two-component signal transduction system RR06/HK06 regulates expression of cbpA in Streptococcus pneumoniae. Proc Natl Acad Sci U S A 102: 7701-7706.
- Sebert ME, Palmer LM, Rosenberg M, Weiser JN (2002) Microarray-based identification of htrA, a Streptococcus pneumoniae gene that is regulated by the CiaRH two-component system and contributes to nasopharyngeal colonization. Infect Immun70: 4059-4067.
- Cheng Q, Campbell EA, Naughton AM, Johnson S, Masure HR (1997) The com locus controls genetic transformation in Streptococcus pneumoniae. MolMicrobiol 23: 683-692.
- Be?ne?dicte Fournier, Andre? Klier (2004) Protein A gene expression is regulated by DNA supercoiling which is modified by the ArlS?ArlR two-component system of Staphylococcus aureus. Microbiology 150: 3807-3819.
- Ulijasz AT, Andes DR, Glasner JD, Weisblum B (2004) Regulation of iron transport in Streptococcus pneumoniae by RitR, an orphan response regulator. J Bacteriol 186: 8123-8136.
- Blue CE, Mitchell TJ (2003) Contribution of a response regulator to the virulence of Streptococcus pneumoniae is strain dependent. Infect Immun 71: 4405-4413.
- Ng WL, Tsui HC, Winkler ME (2005) Regulation of the pspA virulence factor and essential pcsBmurein biosynthetic genes by the phosphorylated VicR (YycF) response regulator in Streptococcus pneumoniae. J Bacteriol 187: 7444-7459.
- Johnsborg O, Eldholm V, H?varstein LS (2007) Natural genetic transformation: prevalence, mechanisms and function. Res Microbiol 158: 767-778.
- Rodriguez AM, Callahan JE, Fawcett P, Ge X, Xu P, et al. (2011) Physiological and molecular characterization of genetic competence in Streptococcus sanguinis. Mol Oral Microbiol 26: 99-116.
- Lindler LE, Macrina FL (1986) Characterization of genetic transformation in Streptococcus mutans by using a novel high-efficiency plasmid marker rescue system. J Bacteriol 166: 658-665.
- Murchison HH, Barrett JF, Cardineau GA, Curtiss R 3rd (1986) Transformation of Streptococcus mutans with chromosomal and shuttle plasmid (pYA629) DNAs. Infect Immun 54: 273-282.
- Perry D, Kuramitsu HK (1981) Genetic transformation of Streptococcus mutans. Infect Immun 32: 1295-1297.
- Tao L, MacAlister TJ, Tanzer JM (1993) Transformation efficiency of EMS-induced mutants of Streptococcus mutans of altered cell shape. J Dent Res 72: 1032-1039.
- van der Ploeg JR (2005) Regulation of bacteriocin production in Streptococcus mutans by the quorum-sensing system required for development of genetic competence. J Bacteriol 187: 3980-3989.
- Claverys JP, Prudhomme M, Mortier-Barriere I, Martin B (2006) Adaptation to the environment: Streptococcuspneumoniae, a paradigm for recombination-mediated genetic plasticity? MolMicrobiol 35: 251-259.
- Prudhomme M, Attaiech L, Sanchez G, Martin B, Claverys JP (2006) Antibiotic stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. Science 313: 89-92.
- Finkel SE, Kolter R (2001) DNA as a nutrient: novel role for bacterial competence gene homologs. J Bacteriol 183: 6288-6293.
- Havarstein LS, Coomaraswamy G, Morrison DA (1995)An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proc Natl Acad Sci USA 92: 11140-11144.
- H?varstein LS (1998) Identification of a competence regulon in Streptococcus pneumoniae by genomic analysis. Trends Microbiol 6: 297-299.
- Kleerebezem M, Quadri LE (2001) Peptide pheromone-dependent regulation of antimicrobial peptide production in Gram-positive bacteria: a case of multicellular behavior. Peptides 22: 1579-1596.
- Martin B, Granadel C, Campo N, H?nard V, Prudhomme M, et al. (2010) Expression and maintenance of ComD-ComE, the two-component signal-transduction system that controls competence of Streptococcus pneumoniae. Mol Microbiol 75: 1513-1528.
- Giammarinaro P, Sicard M, Gasc AM (1999) Genetic and physiological studies of the CiaH-CiaR two-component signal-transducing system involved in cefotaxime resistance and competence of Streptococcus pneumoniae. Microbiology 145: 1859-1869.
- Senadheera DB, Cordova M, Ayala EA, Ch?vez de Paz LE, Singh K, et al. (2012) Regulation of bacteriocin production and cell death by the VicRK signaling system in Streptococcus mutans. J Bacteriol 194: 1307-1316.
- Yamamoto S, Mitobe J, Ishikawa T, Wai SN, Ohnishi M, et al. (2014) Regulation of natural competence by the orphan two-component system sensor kinase ChiS involves a non-canonical transmembrane regulator in Vibriocholerae. Mol Microbiol 91: 326-347.
- Mascher T, Zimmer SL, Smith TA, Helmann JD (2004) Antibiotic-inducible promoter regulated by the cell envelope stress-sensing two-component system LiaRS of Bacillus subtilis. Antimicrob Agents Chemother 48: 2888-2896.
- Klinzing DC, Ishmael N, Dunning Hotopp JC, Tettelin H, Shields KR, et al. (2013) The two-component response regulator LiaR regulates cell wall stress responses, pili expression and virulence in group B Streptococcus. Microbiology 159: 1521-1534.
- Suntharalingam P, Senadheera MD, Mair RW, L?vesque CM, Cvitkovitch DG (2009) The LiaFSR system regulates the cell envelope stress response in Streptococcus mutans. J Bacteriol 191: 2973-2984.
- Reyes J, Panesso D, Tran TT, Mishra NN, Cruz MR, et al. (2015) A liaR Deletion Restores Susceptibility to Daptomycin and Antimicrobial Peptides in Multidrug-Resistant Enterococcus faecalis. J Infect Dis 211:1317-1325.
- Mascher T, Heintz M, Z?hner D, Merai M, Hakenbeck R (2006) The CiaRH System of Streptococcus pneumoniae Prevents Lysis during Stress Induced by Treatment with Cell Wall Inhibitors and by Mutations in pbp2x Involved in beta-Lactam Resistance. J Bacteriol 188: 1959-1968.
- Fritsch F, Mauder N, Williams T, Weiser J, Oberle M, et al. (2011) The cell envelope stress response mediated by the LiaFSRLm three-component system of Listeria monocytogenes is controlled via the phosphatase activity of the bifunctional histidine kinase LiaSLm. Microbiology 157: 373-386.
- Bergholz TM, Tang S, Wiedmann M, Boor KJ (2013) Nisin resistance of Listeria monocytogenesis increased by exposure to salt stress and is mediated via LiaR. Appl Environ Microbiol 79: 5682-5688.