Keywords
Heart failure; HFpEF; HFmrEF; HFrEF; Micro vesicles; Diagnosis; Prediction
Abbreviations
BMI: Body Mass Index; BNP: Brain Natriuretic Peptide; CHF: Chronic Heart Failure; CV: Cardiovascular; EVs: Extracellular Vesicles; HF: Heart Failure; HFmrEF: Chronic HF with Mid-Range Ejection Fraction; HFpEF: Chronic HF with Preserved Ejection Fraction; HFrEF: Chronic HF with Reduced Ejection Fraction; HSP: Heart Shock Protein; GFR: Glomerular Filtration Rate; hs-CRP: High Sensitive C-Reactive Protein; HDL-C: High-Density Lipoprotein Cholesterol; ICAM: Intracellular Adhesion Molecule; LDL-C: Low- Density Lipoprotein Cholesterol; LVEF: Left Ventricular Ejection Fraction; MVs: Micro Vesicles; MI: Myocardial Infarction; STEMI ST: Segment Elevation Myocardial Infarction; PGF: Placental Growth Factor; PLGA: Poly Lactic-co-Glycolic Acid; VCAM: Vascular Cell Adhesion Molecule; VEGF: Vascular Endothelial Growth Factor
Introduction
Although last decades shown the prevalence of heart failure (HF) tend to be decreased in developed countries, morbidity and mortality of this disease remains unacceptably high [1,2]. Additionally, due to a higher incidence of recurrent admission to the hospital global expenditures for HF care are discussed as extremal and exaggerate [2,3]. Current European clinical guideline for HF determines three phenotypes of HF, such as HF with reduced left ventricular ejection fraction (HFrEF), HF with preserved (HFpEF) left ventricular ejection fraction and HF with mid-ranged (HFmrEF) left ventricular ejection fraction, which exhibited increased risk of death and HF-related clinical outcomes including re-hospitalization compared to individuals who did not have HF [1,4]. Keeping in mind HFrEF patients had the worst longterm clinical prognosis, but HFpEF could show better survival rate. In contrast to the patients with HFrEF and HFpEF individuals with HFmrEF generally had intermediate clinical characteristics, but the risk of death was similar HFrEF [5,6]. Interestingly, the survival rate may relate to age, sex, ethnicity, socioeconomic status and co-morbidities in any patients with HF regardless of phenotype of the disease [5,6]. However, clinical outcome in all HF categories and require to be categorized as poor.
The clinical correlates with prognosis sufficiently distinguished each other in patients with various phenotypes of HF and associated with scattered risk marker profile that is explained suggesting differences in underlying pathophysiological mechanisms. For instance, patients with HFrEF were more likely to have male gender, current smoker status, coronary disease, previous myocardial infarction, but female gender, atrial fibrillation, obesity, elevated urinary albumin excretion and cystatin C levels were frequently conferred HFpEF or HFmrEF [7]. However, any phenotypes of HF were determined with increased N-terminal pro-B-type natriuretic peptide (NT-proBNP) [8]. Interestingly, the clinical outcomes in patients with different HF phenotypes have been occurred quite similar and that is important challenge for current medical service [9-11].
Despite improving the HF care remains a priority for health professionals worldwide and novel technologies (cardiac resynchronization therapy, implantable defibrillator/ cardioverters, regenerative cell therapy, personified translational care) are more and more implemented into routine clinical practice, the HF bedevils a devastating impact on patients and prognosis of the disease remains to be poor [10,11].
Although several studies have reported the utilization of numerous biological markers in HF for improving diagnosis, prediction, and therapy, but natriuretic peptides, galectin-3, soluble ST2, and high sensitive cardiac specific troponins were validated only in US and natriuretic peptides are recommended in EU [4,12]. Indeed, both 2012 European Society of Cardiology (ESC) Guidelines for the Diagnosis and Treatment of Acute and Chronic Heart Failure and 2013 American College of Cardiology Foundation/American Heart Association (ACCF/AHA) Guideline for the Management of Heart Failure are positively accepted by many clinicians in different countries as reliable and simple tool for diagnosis and prognosis of suspecting HFrEF, HFmrEF and HFpEF [13,14]. At the same time, individualized risk stratification and personalized medical care remain challenging for many patients with established HF, because current diagnostic and medical strategy based on universal clinical criteria and biomarker approaches [15]. In fact, NPs are universal diagnostic biomarkers, while other biomarkers that were disputed in the clinical guidelines mentioned above including ST-2, galectin-3, growth differentiation factor-15 mid-regional prohormone are emerging as potentially useful prognostic markers in HF. However, there is not strong evidence that all these biomarkers are best fitted for monitoring of response in medical care in HF. Indeed, most of the research on biomarkers in HF has appropriately focused on their use as diagnosis and predictive tools, another emerging role of biomarkers predominantly NPs and galectin-3 is as part of the design of several clinical trials, which are focused on biomarkers as surrogates in HF [16]. Current findings reported that NPs and galectin-3 are biomarkers the concentrations of which have demonstrated a potency to increase depending on advance of HF and to decrease in resulting of euvolemic state achievement. However, high biological variability, depending on age, metabolic comorbidities and kidney function, as well as lowered ability to predict different phenotypes of HF is considered as a serious limitation for serial measures of traditional biomarkers [17].
To note, wide spectrum of novel biological markers reflecting different stages of nature evolution of HF could stratify patients at risk and translating medical care based on individual-derived particularities of the disease including phenotype of HF, etiology, co-morbidities, and complications [18,19]. Additionally, micro vesicles (MVs), which are known as biomarker of vascular injury and endothelial dysfunction appear to be best fitted to personified biomarker stratification at risk of HF developing and HF-related events [20-22]. The aim of the review is to summarize new knowledge for the role of various MPs in diagnosis and prognosis of HF.
Definition, classification, structure and regulation of micro vesicles
Table 1 reports the basic characteristic of sub-population of extracellular vesicles (EVs) with diameter average from 100 to 1000 nm originated from circulating cells. EVs are determined phospholipid-based endogenously produced particles (30- 1000 nm in diameter), which contain cell-specific proteins, glycoproteins, lipids, nucleic acids and active molecules. Ability to secret EVs is proven for numerous cells including circulating blood cells cardiomyocites, mature endothelial cells, immune cells (including T- and B-lymphocytes, dendritic cells), astrocytes, and tumor cells.
| Population of vesicles |
Diameter, nm |
Origin |
Main contained components |
Best characterized cellular sources |
Markers |
| EV |
30-1000 nm |
cell membranes |
regulatory proteins (i.e., heat-shock proteins, tetraspanin), lipids, active molecules, nucleic acids (mRNA, miRNA), cytokines, growth factors, hormones, procoagulant phosphatidylserine, likely complement |
All cell types |
Annexin V binding, tissue factor and cell-specific markers |
| Platelets, RBC and endothelial cells |
| MV |
50–1000 nm |
plasma membranes |
Platelets, RBC and endothelial cells |
| Small-size MPs |
<50 nm |
plasma membranes |
Endothelial cells |
CD133+, CD63- |
| Exosomes |
30–100 nm |
endosomal membranes |
Immune cells and tumors |
CD63, CD61, CD63, CD81, CD9, LAMP1 and TSG101 |
| Ectosomes |
100–350 nm |
plasma membranes |
Platelets, RBC, activated neutrophils, and endothelial cells |
TyA, C1q |
| Late endosomes |
50–1000 nm |
endosomal membranes |
close-packed lumenal vesicles |
Immune cells and tumors |
Annexin V binding, DNA content |
| Apoptotic bodies |
0.5-3.0 µm |
plasma membranes |
Pro-apoptotic molecules, oncogenic receptors |
Cell lines |
Abbreviations: EVs, extracellular vesicles; MPs, micro particles; MV, micro vesicles; RBC, red blood cells.
Table 1: Classification and key features of extracellular vesicles.
There are numerous classifications of EVs depending on origin, sizes, immune phenotypes, and triggers that induce EV realize. For instance, subsets that originated from endosomal membrane are named exosomes, particles that are released on the exocytosis are called late endosomes, EVs released from the plasma membrane are identified as ectosomes, and finally actively secreting EVs as a component of cell membrane are named micro vesicles (MVs). Exosomes have average size from 30 to 100 nm in diameter, the MVs are usually determined as particles with 50 to 1000 nm in diameter, ectosomes have diameter in average from 100 to 350 nm, small-size MVs and apoptotic bodies have diameter 50 nm and less [23-25].
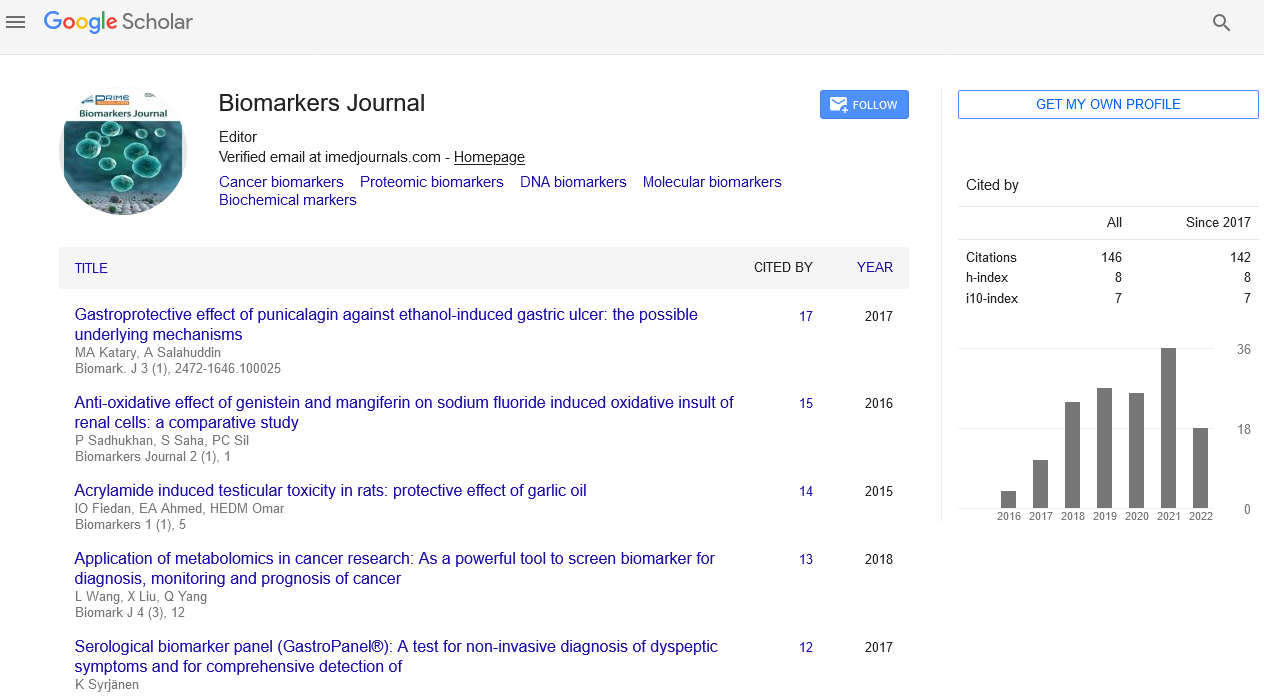
Ninety percent and more of MPs in healthy volunteers are originated from platelets, 10% of MV population is reported as granulocytes-derived particles and about 5% of MCs are reported as endothelial cell-derived MVs, red blood cells (RBCs) and monocytes [26]. Since all types of particles contain surface proteins derived from their cell of origin (including antigen-presenting cells), while there are additional biomarkers confirming origin of the MVs (Figure 1). Table 2 reports the most common features of different MV subsets. It has suggested that the difference between number and immune phenotypes of several subsets of MVs could use as promising biomarker with predictive value of CV diseases including HF.
| Subsets of MVs |
Markers |
The most common methods of detection |
| Derived from resting or activated cells |
| |
Granulocytes |
CD24+CD11c− CD66b/CD66acde |
Flow cytometry western blotting, mass spectrometry, electron microscopic technique, SPRi microscopy |
| Monocytes |
CD14 |
| Microphages |
CD11b+ CD64+/− Ly6Clo |
| Endothelial cells |
CD144, CD62E |
| T cells |
CD4 or CD8 |
| B cells |
CD20 |
| Dendritic cells |
CD1a, CD14, CD141, CD80, CD85, CD86 |
| ICAM(+) cells |
CD54 |
| VCAM(+)cells |
CD106 |
| Platelets |
CD41 and/or CD61 |
| RBCs |
CD235a, CD44, CD47, CD55, CFSE, annexin V and anti-glycophorin A |
| Derived from activated or tumor cells |
Annexin V binding, CD63, CD81, CD9, LAMP1 and TSG101 |
Flow cytometry, capture based assays |
| Derived from apoptotic cells |
Annexin V, DNA content, histones |
Flow cytometry |
Abbreviations: ICAM, intracellular adhesion molecule; VCAM, vascular cell adhesion molecule; SPRi microscopy, nano-particles- surface plasmon resonance - based imaging microscopy; CFSE, carboxyfluorescein diacetate succinimidyl ester.
Table 2: The most common features of MV subsets.
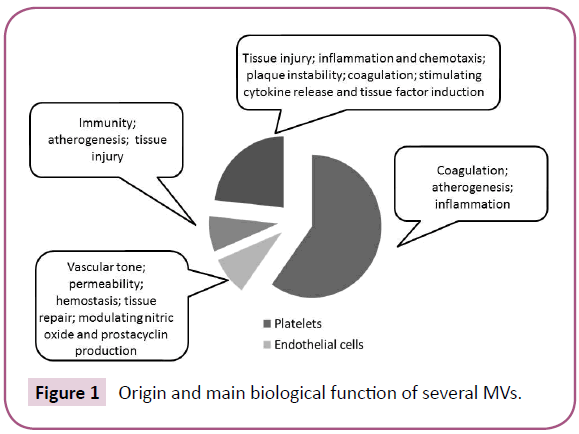
Figure 1: Origin and main biological function of several MVs.
Biological role and function of MPs
MVs have great potentiality in material applications [27], but initially they were found as cell debris and they biological function was not recognized completely. Developing analytic technologies attenuate recognition, determination, and measures of several subsets of MVs [28].
It has been postulated that MPs are a cargo for various molecules including proteins, RNAs, micro-RNAs, DNA fragments, lipids, active molecules, hormones, growth factors. MPs play a central role in cell-to-cell cooperation via transfer these substances from mother cells to recipient cells. Within last decade MPs were found as independent mediator of cell function regardless of cargo. For instance, MC producing with activated cells or apoptotic cells may effect mutually opposite impact of target cells attenuating differentiation and growth or leading to weak function and cell injury [29-31]. Gaining evidence of a pivotal role of MVs originated from different cells (RBCs, mononuclears, endothelial cells, platelets, immune cells) in nature evolution of various diseases including CV diseases, cancer, sepsis, pre- and eclampsia, autoimmune and metabolic states [32-35].
Mononuclear cell-derived MVs are involved in several processes including inflammation, blood coagulation, and thrombosis [36,37]. Mononuclear cells are able to release MVs with proinflammatory capacities as resulting in activation p38 mitogenactivated protein kinase signaling due to effect of several cytokines, bacterial antigens, P-selectin, histamine, catecholamines, angiotensin-II, and cigarette smoking [38-42]. Additionally there is spontaneous secretion of MVs by mononuclear cell without obvious cause in physiological state [42-44].
The most common mechanisms by which MVs may exert their biological functions on target cells are reported (Figure 2).
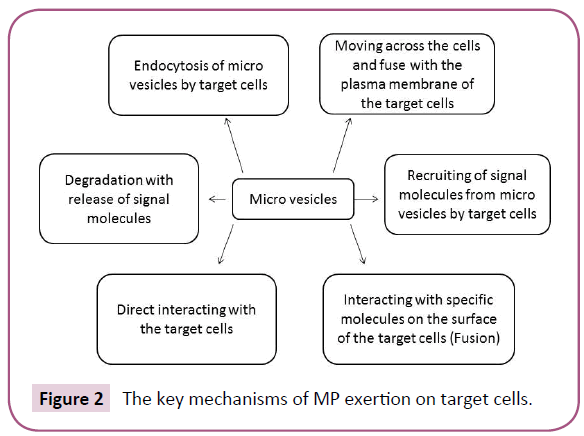
Figure 2: The key mechanisms of MP exertion on target cells.
MVs originated from mononuclears and mature endothelial cells may incorporate in several places of endothelium and damages its integrity directly or via cooperation with oxidized lipids [45- 47]. It is interesting that older individuals demonstrated higher level of pro-coagulant mononuclear cell-derived MVs compared with young people in physiological state and in diseases [48]. The most common triggers for mononuclear cell-derived and endothelial cell-derived MVs were laminar shear stress on endothelium, antigen stimulation, coagulation/thrombosis, endotoxins, activated/apoptotic cells, ischemia/hypoxia and malignancy [49-53].
Finally, mononuclear cell-derived and endothelial cell-derived MVs play an important role in cell-to-cell cooperation, vascular reparation, neovascularization, angiogenesis, oxidative stress, inflammation, and coagulation [53-55].
MVs originated from RBCs act as predominantly trigger of coagulation [56] that has been found in several states including stroke, acute coronary syndrome, myocardial infarction, acute HF, hemolysis, sickle cell disease, thalassemia, chronic kidney disease, trauma, and sepsis [57-62]. Therefore, RBC-derived MPs may be produced ex vivo during cold storage of RBCs because they present factor XI on their surface [63]. All these findings confirm the hypothetical role of RBC-derived MVs in attenuation of coagulation cascade and promoting thrombotic state [64-66]. On the other hand, RBC-derived MVs have fibrinolytic activity predominantly related to plasminogen content on surfaces of them [67].
Koshiar et al. [68] reported that RBC-derived MP surface contains protein C system, which act as natural anticoagulant in vivo. Nevertheless, cell-free RNAs, which are derived from RBCs-MPs, play a pivotal role in transfusion-related immune modulation [56,69].
Platelet-derived MVs are produced due to activation, stress, or apoptosis of platelets like several types of nucleus cells. Because platelet-derived MVs contain phospholipid-based membranes and express in numerous receptors on their surface, they are found as pro-coagulant and complement triggers. The most common biological role of platelet-derived MV is regulating hemostasis, coagulation, inflammation, and probably promoting tissue regeneration and cell repair [70-73].
Table 3 is accumulated information about established role of different subsets of MVs in several diseases. Additionally, Figure 3 is reported the crucial role of MVs in physiological events occurring in tissues and organs.
| Subsets of MVs |
Relation to pathogenic processes |
Relation to diseases |
| Platelet-derived MVs |
Coagulation, inflammatory processes, thrombosis, and malignancy |
ACS, myocardial infarction, HF, tumor progression, systemic lupus erythematosus, vasculitis |
| Leukocytes-derived MVs |
Endothelial dysfunction, , and vascular inflammation |
CV diseases, infections, sepsis, autoimmune disease |
| Endothelial cell-derived MVs |
Endothelial dysfunction, angiogenesis, tumor growth, and increased oxidative stress |
Hypertension, HF, CV diseases, diabetes, metabolic syndrome, obesity, vasculitis, sepsis, inflammation, thrombosis, pulmonary hypertension, acute kidney injury, chronic kidney disease, antiphospholipid syndrome, preeclampsia |
| RBC-derived MVs |
Immunomodulation, coagulation, inflammation |
Thrombosis, vasculitis, sepsis, ACS, myocardial infarction, autoimmune disease, chronic kidney disease, antiphospholipid syndrome, preeclampsia, graft rejection |
| Immune cells-derived MPs |
Cargo of tumor rejection antigens |
Tumor progression |
Abbreviations: MVs, micro vesicles; CV, cardiovascular disease; ACS, acute coronary syndrome; RBC, red blood cells.
Table 3: The critical role of MVs in pathogenesis of several diseases.
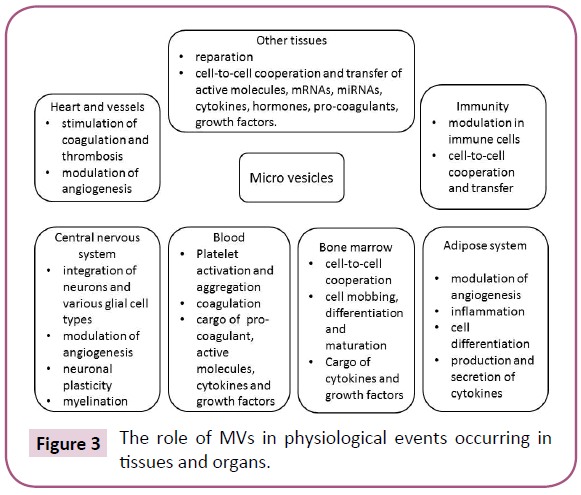
Figure 3: The role of MVs in physiological events occurring in tissues and organs.
Platelet-derived MVs were found a cargo tool of bioactive molecules including growth factors, signaling molecules and fragments of chromatin [74].
Endothelial cells-derived MVs are produced by several triggers such as angiotensin II, lipopolysaccharide substances, and hydrogen peroxide [49]. Because all these factors are central players in the pathogenesis of CV diseases, it is not surprising that endothelial cells-derived MVs are involved in the target organ damage, either exacerbating disease progression or triggering a repair response [75-79]. Noted that endothelial cells-derived MVs (EMVs) originated from apoptotic cells had chromatin substance and may destroy target cell directly, promote coagulation/ thrombosis [75,76]. In contrast, EMVs derived from activated mature endothelial cells do not contain chromatin components and have angiogenic properties contributing to tissue repair [77-80]. In this context, origin of endothelial cells-derived MVs might have the crucial for vascular integrity and supporting endothelial function. All these are recognized as an important mechanisms of developing CV disease and HF for which endothelial dysfunction is key element in pathogenesis of the diseases.
Measure of MVs
There are several controversies in point of view on methodology of assay of morphology, transcriptomics, and proteomics of circulating MVs [81,82]. Indeed, there is sufficient difficulty in separating and isolation of MVs. All these depend on the limits of detection of conventional flow cytometry, and other analytical methods that enable determine of MVs in biological fluids and recognize of their origin [83]. Therefore, the majority of methods of MV detection remain costly and time consuming [84].
The conventional approach for measuring the MVs is based on commonly used flow cytometry, mass spectrometry-based proteomic methods, Western blot analysis, electron and atomic microscopy, and nano-particle tracking analysis (NTA) [85]. All of them demonstrate serious limitations in sensitivity, specificity, probability and accuracy in resolution of MVs, while commercial flow cytometry technique remains gold standard at this way [86,87].
Flow cytometry is a well-standardized and accepted method for cell identification and detection, although the standard tool requires special attention when measuring MVs in diameter less 200 nm [88]. Therefore, there is serious limitation regarding ability to recognized small-size particles like MVs in diameter, i.e. low-density lipoproteins. However, the flow cytometer provides the possibility to measure MPs directly in plasma samples and to analyze MV-subsets [89].
NTA technique is commercially available method that allows visualizing MV and measuring number of them, while detection limit for this method is 50 nm. Western blot analysis, atomic and electron microscopy require high qualified team with great experience in this field and are much expensive. To overcome size limit highly sensitive fluorescent (HSF) microscopy and advanced flow cytometry technique based on BD Horizon Violet Proliferation dye are used now as alternative techniques. However, two and more consequently performing analytic techniques appear to be much more promising in detection of wide-range of MVs [86,87]. Probably nano-particles- surface plasmon resonance-based imaging microscopy (SPRi microscopy) could be simple and affordable method for MV determining in future [88-90]. However, SPR signal between cell edges and substratum sufficiently improves identification of MV edges and segmentation of MV areas [91]. Probably simultaneous utilization of a HSF microscopy and SPRi microscopy could enhance the sensitivity and selectivity of novel biosensor platform besides in detection of small sized MVs [92-96].
Additionally, there are not commercially available techniques that could be promising in MV detection, such as surface-assisted laser desorption/ionization mass spectrometry (SALDI-MS) Raman micro-spectroscopy, micro nuclear magnetic resonance technique, small-angle X-ray scattering, and anomalous smallangle X-ray scattering [97,98]. The main advantage of these approaches is detection of MVs isolated from multiple biological sources without previous sample preparation per protocols. There is integrated system entitled highly sensitive magnetic resonance imaging that allows detecting MV antigens with microfluidic chip and labelling target-specific magnetic nanoparticles [99]. Thus innovative approach may differentiate MVs derived from tumor cells from non-tumor host cells [99].
There are commercial platforms that offer simultaneous, labelfree optical biological sensing of MVs with affordable integrated analytic circuits [100]. Despite these successes, isolation, purification and content analysis of MVs remain to be failed to use in routine laboratory practice [101]. Finally, any single method of MV determination does not appear to be superior a combined methods. Novel models and techniques for identification of MVs based on real-time and label-free biosensors, such as SPR, demonstrate advantages before conventional old methods.
MVs in Cardiovascular Disease
Number of circulating MVs originated from several cells is changed in patients with known CV disease including HFrEF, HFmrEF and HFpEF [102,103]. Recent studies have shown elevated levels of MVs originated from numerous blood cells MVs was found in the patients with established CV disease including HF compared with healthy volunteers [102-107]. However, there are several controversies about the role of MVs originated from various blood cells in HF.
RBC-derived MPs
Previous evidence has addressed to the role of erythrocytederived MVs (RBC-derived MVs) in coagulation, inflammation, and immunity, but later some suggestions about direct involvement of RBC-derived MVs in the pathogenesis of HF begin to appear. Indeed RBC-derived MVs are mobilized from gaining thrombi into the distal blood flow in STEMI and may be detected in peripheral blood [108]. Probably, newly and ongoing thrombosis could determine via measure of pattern of RBCs-derived MVs [108,109].
Sansone et al. [110] reported that there were elevated levels of RBC-derived MVs, WBC-derived MVs endothelial cell-derived MVs and platelet-derived MPs in patients after implantation of left ventricular mechanical support devices. Authors suspected that mechanical supporting can be a causing factor leading to this phenomenon. In contrast, there is evidence that there was no significant difference between number of RBC (CD235a+)- derived MVs in patients with sudden cardiac death due to STEMI and individuals with stable coronary artery disease [111].
However, numerous findings confirm the causality role of elevated RBCs-derived MVs in CV events resulting in blood transfer, hemoglobinopathies, transplant rejection and autoimmune conflict. Unfortunately, causative role of RBCs-derived MVs in CV disease and HF in not established completely.
White blood cells-derived MVs
The white blood cells-derived (WBCs) MPs were found in higher concentrations in the patients with acute and chronic coronary syndromes, HF and metabolic disease including T2DM, abdominal obesity [112-114]. There is evidence that several immune phenotypes like CD14(+) of WBCs-derived MVs were involved in the reparative vascular response to injury in acute coronary syndromes [113]. Morel et al. [114] found that primary PCI treated patients with STEMI had significantly higher levels of leukocytederived CD11a(+) MVs, endothelial cell-derived CD105(+) MVs, and tissue factor-bearing MVs than those who did not treat with PCI [114]. Thus, elevated WBC-derived MVs suggested their pathophysiological role in coronary occlusion in STEMI. Therefore, there is evidence that generation of pro-coagulant mononuclear cells-derived MVs might be under control of leptin, visfatin and other metabolic triggers [115]. Overall, WBC-derived MVs are a marker of cell injury, inflammation and coagulation. Whether these phenotypes of MVs are promising indicator of a risk in HF is not yet clear ane requires to be investigated in large clinical studies.
Platelet-derived MVs
Platelet-derived MVs were found in higher concentrations in numerous diseases including acute coronary syndrome/ myocardial infarction, HF, heparin-induced thrombocytopenia, thrombotic thrombocytopenic purpura, pulmonary thromboembolism, and hemolytic uremic syndrome. At the same time, there was not determined a significant difference between levels of platelet-derived MVs in healthy volunteers and patients with known abdominal obesity, T2DM, antiphospholipid syndrome, infection disease or sepsis [116]. Probably, platelets before secretion of MVs require to be activated with of adhered WBCs or endothelial cells [117].
Although the importance of platelet-derived MVs in HF pathogenesis remains unclear [116-119], there is evidence that platelet-derived MVs attenuate shaping foam cells in atherosclerotic plaque and thereby may contribute plaque rupture and acute coronary syndrome, which are known predictors of HF [120]. However, some fractions of MVs derived from platelets expressed P-selectin and/or CD63 and play a pivotal role in thrombus formation and atherothrombotic events [121,122]. Michelsen et al. [123] reported that the elevated level of platelet-derived MPs in survivors of acute myocardial infarction associated strongly with thrombosis and soluble CD40 ligand determined in peripheral blood. Probably, MVs derived from platelets are a factor, which directly mediates endothelial dysfunction and accelerate atherosclerosis, while plateletderived MVs may cooperate with endothelial progenitor cells in vascular reparation [116,120,122-124]. Indeed, platelet-derived MVs are able to interact with early outgrowth cells and prevent vascular injury with modulating regenerative ability of progenitor cells [125,126]. Baj-Krzyworzeka et al. [127] shown that plateletderived MVs attenuated biological functions of hematopoietic cells that potentiated intercellular cross-talk in hematopoiesis and vascular repair. Ohtsuka et al. [128] reported that MVs originated from platelets improved endothelialization of capillary with involving angiogenic progenitor cells into regeneration places. Thus, platelet-derived MVs are promising biological marker for vascular dysfunction and CV events that may be incorporated into program of risk stratification in patients with known CV diseases and probably HF [129].
Endothelial cells-derived MVs
There is large body of evidence that the number of endothelial cells-derived MVs (EMVs) may be a marker of endothelial dysfunction with predictive value for CV events and diseases. Recent clinical studies have shown that number of CD31+/ annexin V+ EMVs associated with altered endothelial function and clinical outcomes in patients with stable coronary artery disease [130,131]. Additionally, Huang et al. [132] reported that both increased number of circulating CD31+/annexin V+ EMVs and lowered number circulating EPCs are able to predict a CV risk in hypertension. The increased number EMVs in peripheral blood were found in individuals with acute coronary syndrome and it correlated well with a risk of sudden cardiac death [112]. Nevertheless, a number of EMP with immune phenotype CD42- CD31+ inversely associated with microvascular obstruction grading in acute myocardial infarction [133]. Additionally, impaired ratio between number of apoptotic EMVs and EMVs originated from activated mature endothelial cells was elucidated in numerous patients with known HF, established CV disease as well as in patients at high risk of CV disease and events [124,134,135]. This finding has now recognized a marker of vascular dysfunction due to several reasons including inflammation, thrombosis, atherosclerosis, and contributed to a risk CV complications [135-138].
Figure 4 reports the basic mechanisms shaped “impaired immune phenotype” of the cells, which secret MVs in peripheral blood. It seems to be that not just epigenetically transformed mature endothelial cells are able to release of MVs, but progenitor endothelial cells can have similar capability also. There is suggestion that mature endothelial cells can be under various epigenetic stimuli (ischemia/hypoxia, increased glucose level, inflammatory cytokine and lipid abnormalities) and they turn into “functionally incompetence” associated with worsening secretom. All these lead to altered signaling between mature cells and progenitor cells and thereby ascertain a mechanism of shaping altered vascular reparation, which in not able counteract microvasculature damages [139-145].
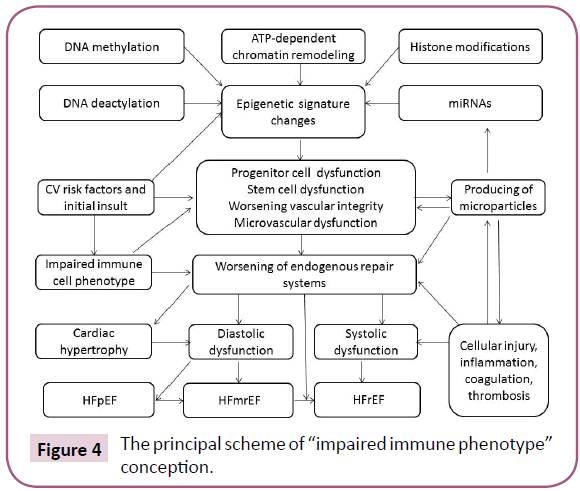
Figure 4: The principal scheme of “impaired immune phenotype” conception.
Thus, in HF pattern of EMVs coordinates an impact of conventional CV risk factors (i.e. insulin resistance, inflammation, thyroid dysfunction, T2DM, abdominal obesity) on vascular reparation and thereby plays a pivotal role in developing endothelial dysfunction that is a central player in HF advance [146-148].
Diagnostic and predictive value of circulating MVs in several phenotypes of HF
Signature of circulating MVs reflecting various stage of pathogenesis of HF could be a biomarker of a risk and severity of HFrEF/HFmrEF and probably HFpEF [20]. Bank et al. [149] reported that both MVs-counts and MVs-content may be novel biomarkers with diagnostic and predictive values for HF. Recently numerous MVs with different origin are detecting to improve conventional risk scores of HF, whereas number and immune phenotypes of EMVs appear to be more promising today. There is evidence that elevated levels of CD62+ MVs, which are secreted with activated mature endothelial cells, may elucidate rather endothelial dysfunction, but lowered number of CD62+ MVs is reported as a marker of severe endothelium injury [21,150].
The concept of “Impaired” Immune phenotype of circulating EMVs as HF biomarker
Altered ratio between numbers of circulating EMVs with different origin is known as impaired phenotype phenomenon. Recently it was found in individuals with endothelial dysfunction and it correlated well with severity of vascular damage [151-154]. Some studies confirmed the idea that simple signature of MVs does not correspond to the number of CV risk factors and that circulating number of single MVs does not adequately elucidate a risk of vascular event and vascular complications [144,155]. The number of apoptotic EMVs alone and adjusted to number of mononuclear progenitor cells shown a much more probability and discriminate value for HF clinical outcomes compared to NT-proBNP, soluble ST2 and galectin-3 used alone [156,157]. All these data led to new predictive score based on serial measure of circulating biomarkers including EMVs, NT-proBNP, soluble ST2 and galectin-3 [158,159]. Thus, “impaired immune phenotype” of circulating EMVs as well as increased number of apoptotic cellderived MVs are novel biomarkers of HF with possible predictive capabilities.
Conclusion
There is gaining interest in the scientific community toward the role of MVs in pathogenesis of HF. MPs are discussed a mediator of several biological processes affecting growth, differentiation, and proliferation of tissues, as well as MVs are essential for immune response, regulation of angiogenesis, neovascularization, and coagulation. There is large number of evidence that number and immune phenotypes of circulating MVs originated from numerous circulating cells (erythrocytes, monocytes, platelets, endothelial cells and tumor cells) could be markers of severity of vascular injury, and endothelial dysfunction. Moreover, there are data received from clinical trials that confirm a participation of EMVs in vascular reparation. Therefore, altered immune phenotype of circulating EMVs associated with increased number of apoptotic cell-derived MVs was found a predictor of CV events and CV disease in patients with HF. Although EMVs are promising biomarkers in HFrEF, HFmrEF and HFpEF, large clinical trials are needed to be confirmed the probability of number and altered immune phenotype EMVs in HF.
Acknowledgments
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Funding and Grants
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of Interests
Not declared
References
- Riet VEE, Hoes AW, Wagenaar KP, Limburg A, Landman MA, et al. (2016) Epidemiology of heart failure: the prevalence of heart failure and ventricular dysfunction in older adults over time. A systematic review. Eur J Heart Fail 18: 242-252.
- Jorge AL, Rosa ML, Martins WA, Correia DM, Fernandes LC, et al. (2016) The prevalence of stages of heart failure in primary care: a population-based study. J Card Fail 22: 153-157.
- Banerjee P (2016) Heart failure with preserved ejection fraction: A clinical crisis. Int J Cardiol 204: 198-199.
- Yancy CW, Jessup M, Bozkurt B, Butler J, Casey DE, et al. (2013) ACCF/AHA guideline for the management of heart failure: A report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines. J Am Coll Cardiol 62: e147-e239.
- Buja A, Solinas G, Visca M, Federico B, Gini R, et al. (2016) Prevalence of heart failure and adherence to process indicators: which socio-demographic determinants are involved. Int J Environ Res Public Health 13: 238.
- Kosiborod M, Lichtman JH, Heidenreich PA, Normand SL, Wang Y, et al. (2006) National trends in outcomes among elderly patients with heart failure. Am J Med 119: e1-e7.
- Frankenstein L, Clark A.L, Ribeiro JP (2012) Influence of sex on treatment and outcome in chronic heart failure. Cardiovasc Ther 30: 182-192.
- Rathore SS, Masoudi FA, Wang Y, Curtis JP, Foody JM, et al. (2006) Socioeconomic status, treatment, and outcomes among elderly patients hospitalized with heart failure: findings from the national heart failure project. Am Heart J 152: 371-378.
- Brouwers FP, de Boer RA, van der Harst P, Voors AA, Gansevoort RT, et al. (2013) Incidence and epidemiology of new onset heart failure with preserved vs. reduced ejection fraction in a community-based cohort: 11-year follow-up of PREVEND. Eur Heart J 34: 1424-1431.
- Hawkins NM, Virani SA, Sperrin M, Buchan IE, McMurray JJ, et al. (2015) Predicting heart failure decompensation using cardiac implantable electronic devices: a review of practices and challenges. Eur J Heart Fail.
- McMurray JJ (2015) Improving outcomes in heart failure: a personal perspective. Eur Heart J 36: 3467-3470.
- McMurray JJ, Adamopoulos S, Anker SD, Auricchio A, Bohm M, et al. (2012) ESC guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: The task force for the diagnosis and treatment of acute and chronic heart failure 2012 of the European society of cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur J Heart Fail 14: 803-869.
- Butler J, Fonarow GC, Zile MR, Lam CS, Roessig L, et al. (2014) Developing therapies for heart failure with preserved ejection fraction: current state and future directions. JACC Heart Fail 2: 97-112.
- D'Elia E, Vaduganathan M, Gori M, Gavazzi A, Butler J, et al. (2015) Role of biomarkers in cardiac structure phenotyping in heart failure with preserved ejection fraction: critical appraisal and practical use. Eur J Heart Fail.
- Chamberlain AM, St Sauver JL, Gerber Y, Manemann SM, Boyd CM, et al. (2015) Multimorbidity in heart failure: a community perspective. Am J Med 128: 38-45.
- Berezin A (2018) Circulating biomarkers in heart failure: diagnostic and prognostic importance. J Lab Precis Med 3: 36.
- Berezin A (2017) Can biomarkers be useful for cardiovascular risk assessment in patients with different phenotypes of obesity? J Basic Clin Pharma 8: S01?S05.
- Paulus WJ, Tschöpe CA (2013) novel paradigm for heart failure with preserved ejection fraction: comorbidities drive myocardial dysfunction and remodeling through coronary microvascular endothelial inflammation. J Am Coll Cardiol 62: 263-271.
- Fujisue K, Sugiyama S, Matsuzawa Y, Akiyama E, Sugamura K, et al. (2015) Prognostic significance of peripheral microvascular endothelial dysfunction in heart failure with reduced left ventricular ejection fraction. Circ J 79: 2623-2631.
- Dignat-George F, Boulanger CM (2011) The many faces of endothelial microparticles. Aterioscler Thromb Vasc Biol 31: 27-33.
- Burger D, Touyz RM (2012) Cellular biomarkers of endothelial health: microparticles, endothelial progenitor cells and circulating endothelial cells. J Am Soc Hypertens 6: 85-99.
- Berezin A (2015) Endothelial derived micro particles: biomarkers for heart failure diagnosis and management. J Clin Trial Cardiol 2: 1-3.
- Cocucci E, Meldolesi J (2015) Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol 25: 364-372.
- Colombo M, Raposo G, Théry C (2014) Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol 30: 255-289.
- Piccin A, Murphy WG, Smith OP (2007) Circulating microparticles: pathophysiology and clinical implications. Blood Rev 21: 157-171.
- Tesselaar ME, Romijn FP, Van DL, Prins FA, Bertina RM, et al. (2007) Micro particle-associated tissue factor activity: a link between cancer and thrombosis? J Thromb Haemost 5: 520-527.
- Sarkar S, Dasgupta AK (2015) Microparticle of drug and nanoparticle: a biosynthetic route. Pharmacol Res Perspect 3: e00188
- Gu Z, Jing C, Ying YL, He P, Long YT (2015) In situ high throughput scattering light analysis of single plasmonic nanoparticles in living cells. Theranostics 5: 188-195.
- Gong J, Jaiswal R, Dalla P, Luk F, Bebawy M (2015) Microparticles in cancer: A review of recent developments and the potential for clinical application. Semin Cell Dev Biol 40: 35-40.
- Das S, Halushka MK (2015) Extracellular vesicle microRNA transfer in cardiovascular disease. Cardiovasc Pathol 24: 199-206.
- Berezin A, Zulli A, Kerrigan S, Petrovic D, Kruzliak P (2015) Predictive role of circulating endothelial-derived microparticles in cardiovascular diseases. Clin Biochem 48: 562-568.
- Jadli A, Sharma N, Damania K, Satoskar P, Bansal V, et al. (2015) Promising prognostic markers of Preeclampsia: New avenues in waiting. Thromb Respii 136: 189-195.
- Greening DW, Gopal SK, Mathias RA, Liu L, Sheng J, et al. (2015) Emerging roles of exosomes during epithelial-mesenchymal transition and cancer progression. Semin Cell Dev Biol 40: 60-71.
- Martinez MC, Andriantsitohaina R (2011) Microparticles in angiogenesis: therapeutic potential. Circ Res 109: 110-119.
- Souza AC, Yuen PS, Star RA (2015) Microparticles: markers and mediators of sepsis-induced microvascular dysfunction, immunosuppression, and AKI. Kidney Int 87: 1100-1108.
- Neri T, Pergoli L, Petrini S, Gravendonk L, Balia C, et al. (2016)Particulate matter induces prothrombotic microparticle shedding by human mononuclear and endothelial cells. Toxicol In Vitro 32: 333-338.
- Aleman MM, Gardiner C, Harrison P, Wolberg AS (2011) Differential contributions of monocyte- and platelet-derived microparticles towards thrombin generation and fibrin formation and stability. J Thromb Haemost 9: 2251-2261.
- Cordazzo C, Petrini S, Neri T, Lombardi S, Carmazzi Y, et al. (2014) Rapid shedding of proinflammatory microparticles by human mononuclear cells exposed to cigarette smoke is dependent on Ca2+ mobilization. Inflamm Res 63: 539-547.
- Li M, Yu D, Williams KJ, Liu ML (2010) Tobacco smoke induces the generation of procoagulant microvesicles from human monocytes/macrophages. Arterioscler Thromb Vasc Biol 30: 1818-1824.
- Novelli F, Neri T, Tavanti L, Armani C, Noce C, et al. (2014) Procoagulant, tissue factor-bearing microparticles in bronchoalveolar lavage of interstitial lung disease patients: an observational study. PLoS One 9: e95013.
- Vatsyayan R, Kothari H, Pendurthi UR, Rao LV (2013) 4-Hydroxy-2-nonenal enhances tissue factor activity in human monocytic cells via p38 mitogen-activated protein kinase activation-dependent phosphatidylserine exposure. Arterioscler Thromb Vasc Biol 33: 1601-1611.
- Satta N, Toti F, Feugeas O, Bohbot A, Dachary-Prigent J, et al. (1994) Monocyte vesiculation is a possible mechanism for dissemination of membrane-associated procoagulant activities and adhesion molecules after stimulation by lipopolysaccharide. J Immunol 153: 3245-3255.
- Morel N, Morel O, Delabranche X, Jesel L, Sztark F, et al. (2006) Microparticles during sepsis and trauma. A link between inflammation and thrombotic processes. Ann Fr Anesth Reanim 25: 955-966.
- Chung SM, Bae ON, Lim KM, Noh JY, Lee MY, et al. (2007) Lysophosphatidic acid induces thrombogenic activity through phosphatidylserine exposure and procoagulant microvesicle generation in human erythrocytes. Arterioscler Thromb Vasc Biol27: 414-421.
- Kawata J, Aoki M, Ishimaru Y, Ono T, Sagara K, et al. (2015) Mechanism of tissue factor production by monocytes stimulated with neutrophil elastase. Blood Cells Mol Dis 54: 206-209.
- Neri T, Cordazzo C, Carmazzi Y, Petrini S, Balìa C, et al. (2012) Effects of peroxisome proliferator-activated receptor-γ agonists on the generation of microparticles by monocytes/macrophages. Cardiovasc Res 94: 537-544.
- Cordazzo C, Neri T, Petrini S, Lombardi S, Balìa C, et al. (2013) Angiotensin II induces the generation of procoagulant microparticles by human mononuclear cells via an angiotensin type 2 receptor-mediated pathway. Thromb Res 131: e168-e174.
- Forest A, Pautas E, Ray P, Bonnet D, Verny M, et al. (2010) Circulating microparticles and procoagulant activity in elderly patients. J Gerontol A Biol Sci Med Sci 65: 414-420.
- Lynch SF, Ludlam CA (2007) Plasma microparticles and vascular disorders. Br J Haematol 137: 36-48.
- Sabatier F, Camoin-Jau L, Anfosso F, Sampol J, Dignat-George F (2009) Circulating endothelial cells, microparticles and progenitors: key players towards the definition of vascular competence. J Cell Mol Med 13:454-471.
- Burnier L, Fontana P, Kwak BR, Angelillo-Scherrer A (2009) Cell-derived microparticles in haemostasis and vascular medicine. Thromb Haemost 101: 439-451.
- Brodsky SV, Zhang F, Nasjletti A, Goligorsky MS (2004) Endothelium-derived microparticles impair endothelial function in vitro. Am J Physiol Heart Circ Physiol 286: 1910-1915.
- Boulanger CM, Amabile N, Guérin AP, Pannier B, Leroyer AS, et al. (2007) In vivo shear stress determines circulating levels of endothelial microparticles in end-stage renal disease. Hypertension 49: 902-908.
- Buzas EI, György B, Nagy G, Falus A, Gay S (2014) Emerging role of extracellular vesicles in inflammatory diseases. Nat Rev Rheumatol 10: 356-364.
- Vader P, Breakefield XO, Wood MJ (2014) Extracellular vesicles: emerging targets for cancer therapy. Trends Mol Med 20: 385-393.
- Muszynski JA, Bale J, Nateri J, Nicol K, Wang Y, et al. (2015) Supernatants from stored red blood cell (RBC) units, but not RBC-derived microvesicles, suppress monocyte function in vitro. Transfusion 55: 1937-1945.
- Cognasse F, Hamzeh-Cognasse H, Laradi S, Chou ML, Seghatchian J, et al. (2015)The role of microparticles in inflammation and transfusion: A concise review. Transfus Apher Sci 53: 159-167.
- Chakrabarti A, Halder S, Karmakar S (2016) Erythrocyte and platelet proteomics in hematological disorders. Proteomics Clin Appl 10: 403-414.
- Piccin A, Murphy C, Eakins E, Kunde J, Corvetta D, et al. (2015) Circulating microparticles, protein C, free protein S and endothelial vascular markers in children with sickle cell anaemia. J Extracell Vesicles 4: 28414.
- Gao C, Ji S, Dong W, Qi Y, Song W, et al. (2015) Indolic uremic solutes enhance procoagulant activity of red blood cells through phosphatidylserine exposure and microparticle release. Toxins (Basel) 7: 4390-4403.
- Zhang Y, Meng H, Ma R, He Z, Wu X, et al. (2016) Circulating microparticles, blood cells, and endothelium induce procoagulant activity in sepsis through phosphatidylserine exposure. Shock 45: 299-307.
- Harms A, Fuehner T, Warnecke G, Haverich A, Gottlieb J, et al. (2015) Epithelial and erythrocyte microvesicles from bronchoalveolar lavage fluid are elevated and associated with outcome in chronic lung allograft dysfunction. Transplantation 99: 2394-2400.
- Prudent M, Crettaz D, Delobel J, Seghatchian J, Tissot JD, et al. (2015) Differences between calcium-stimulated and storage-induced erythrocyte-derived microvesicles. Transfus Apher Sci 53: 153-158.
- Lacroix R, Plawinski L, Robert S, Doeuvre L, Sabatier F, et al. (2012) Leukocyte- and endothelial-derived microparticles: a circulating source for fibrinolysis. Haematologica 97: 1864-1872.
- van Beers EJ, Schaap MC, Berckmans RJ, Nieuwland R, Sturk A, et al. (2009) CURAMA study group. Circulating erythrocyte-derived microparticles are associated with coagulation activation in sickle cell disease. Haematologica 94: 1513-1519.
- Karpman D, Ståhl AL, Arvidsson I, Johansson K, Loos S, et al. (2015) Complement interactions with blood cells, endothelial cells and microvesicles in thrombotic and inflammatory conditions. Adv Exp Med Biol 865: 19-42.
- Levin G, Sukhareva E, Lavrentieva A (2016) Impact of microparticles derived from erythrocytes on fibrinolysis. J Thromb Thrombolysis 41: 452-458.
- Koshiar RL, Somajo S, Norström E, Dahlbäck B (2014) Erythrocyte-derived microparticles supporting activated protein C-mediated regulation of blood coagulation. PLoS One 9: e104-200.
- Carmeliet P, Jain RK (2011) Molecular mechanisms and clinical applications of angiogenesis. Nature 473: 298-307.
- Goubran HA, Burnouf T, Stakiw J, Seghatchian J (2015) Platelet microparticle: a sensitive physiological "fine tuning" balancing factor in health and disease. Transfus Apher Sci 52: 12-28.
- Sinauridze E, Kireev DA, Popenko NY, Pichugin AV, Panteleev MA, et al. (2007) Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb Haemost 97: 425-434.
- Nomura S (2009) Dynamic role of microparticles in type 2 diabetes mellitus. Curr Diabetes Rev 5: 245-251.
- Varon D, Shai E (2009) Role of platelet-derived microparticles in angiogenesis and tumor progression. Discov Med 8: 237-241.
- Nomura S (2001) Function and clinical significance of platelet-derived microparticles. Int J Hematol 74: 397-404.
- Puddu P, Puddu GM, Cravero E, Muscari S, Muscari A (2010) The involvement of circulating microparticles in inflammation, coagulation and cardiovascular diseases. Can J Cardiol 26: 140-145.
- Jimenez JJ, Jy W, Mauro LM, Soderland C, Horstman LL, et al. (2003) Endothelial cells release phenotypically and quantitatively distinct microparticles in activation and apoptosis. Throm Res 109: 175-180.
- Arteaga RB, Chirinos JA, Soriano AO, Jy W, Horstman L, et al. (2006) Endothelial microparticles and platelet and leukocyte activation in patients with the metabolic syndrome. Am J Cardiology 98: 70-74.
- Quesenberry PJ, Aliotta JM (2010) Cellular phenotype switching and microvesicles. Adv Drug Deliv Rev 62: 1141-1148.
- Berezin AE (2016) Metabolic memory phenomenon in diabetes mellitus: achieving and perspectives. Diab Metab Syndrome Clin Res Rev.
- Inal JM, Kosgodage U, Azam S, Stratton D, Antwi-Baffour S, et al. (2013) Blood/plasma secretome and microvesicles. Biochem Biophys Acta 1834: 2317-2325.
- Choi DS, Kim DK, Kim YK, Gho YS (2013) Proteomics, transcriptomics and lipidomics of exosomes and ectosomes. Proteomics 13: 1554-1571.
- Dinkla S, Brock R, Joosten I, Bosman GJ (2013) Gateway to understanding microparticles: standardized isolation and identification of plasma membrane-derived vesicles. Nanomed 8: 1657-1668.
- Patil R, Ghosh K, Shetty SA (2015) simple clot based assay for detection of procoagulant cell-derived microparticles. Clin Chem Lab Med.
- Niccolai E, Squatrito D, Emmi G, Silvestri E, Emmi L, et al. (2015) A new cytofluorimetric approach to evaluate the circulating microparticles in subjects with antiphospholipid antibodies. Thromb Res 136:1252-1258.
- Shantsila E, Montoro-García S, Gallego P, Lip GY (2014) Circulating microparticles: challenges and perspectives of flow cytometric assessment. Thromb Haemost 111: 1009-1014.
- Orozco AF, Lewis DE (2010) Flow cytometric analysis of circulating microparticles in plasma. Cytometry A 77: 502-514.
- Jy W, Horstman LL, Jimenez JJ, Ahn YS, Biro E, et al. (2004) Measuring circulating cell-derived microparticles. J Thromb Haemost 2: 1842-1851.
- Gardiner C, Ferreira YJ, Dragovic RA, Redman CW, Sargent IL (2013) Extracellular vesicle sizing and enumeration by nanoparticle tracking analysis. J Extracell Vesicles 2: 19671.
- Peterson AW, Halter M, Tona A, Plant AL (2014) High resolution surface plasmon resonance imaging for single cells. BMC Cell Biol 15: 35.
- Peterson AW, Halter M, Tona A, Bhadriraju K, Plant AL (2010) Using surface plasmon resonance imaging to probe dynamic interactions between cells and extracellular matrix. Cytometry A 77: 895-903.
- Robelek R, Wegener J (2010) Label-free and time-resolved measurements of cell volume changes by surface plasmon resonance (SPR) spectroscopy. Biosens Bioelectron 25: 1221-1224.
- Shpacovitch V (2012) Application of Surface Plasmon Resonance (SPR) for the detection of single viruses and single biological Nano-objects. J Bacteriol Parasitol 3: e110.
- Zybin A, Kuritsyn YA, Gurevich EL, Temchura VV, Überla K (2010) Real-time detection of single immobilized nanoparticles by surface plasmon resonance imaging. Plasmonics 5: 31-35.
- Berezin AE (2015) The biosensing of microparticles: benefits and perspectives. ARC J Diab Endocrinol 1: 31-34.
- Berezin AE (2015) Impaired phenotype of circulating endothelial-derived microparticles: novel marker of cardiovascular risk. J Cardiol Ther 2: 273-278.
- Van der Pol E, Coumans F, Varga Z, Krumrey M, Nieuwland R (2013) Innovation in detection of microparticles and exosomes. J Thromb Haemost11: 36-45.
- Guinan TM, Kirkbride P, Della Vedova CB, Kershaw SG, Kobus H, et al. (2015) Direct detection of illicit drugs from biological fluids by desorption/ionization mass spectrometry with nanoporous silicon microparticles. Analyst.
- Shao HL, Chung J, Balaj L, Charest A, Bigner DD, et al. (2012) Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nature Med 18: 18-35.
- Laborde C, Pittino F, Verhoeven HA, Lemay SG, Selmi L, et al. (2015) Real-time imaging of microparticles and living cells with CMOS nanocapacitor arrays. Nat Nanotechnol 10: 791-795.
- Berezin AE (2015) The development of biological molecular sensing techniques to detect microparticles: focus on clinical medicine benefits. J Microb Biochem Technol 7: 236-237.
- Berezin A, Kremzer A (2015) The predictive role of circulating microparticles in patients with chronic heart failure. J Extracell Vesicles 4: 14.
- Berezin AE (2015) Impaired pattern of endothelial derived microparticles in heart failure patients. J Mol Genet Med 9: 1.
- Berezin AE, Kremzer AA, Martovitskaya Yu V, Samura TA, Berezina TA (2015) The association of subclinical hypothyroidism and pattern of circulating endothelial-derived microparticles among chronic heart failure patients. Res Cardiovasc Med 4: e290-294.
- Paudel KR, Panth N, Kim DW (2016) Circulating endothelial microparticles: a key hallmark of atherosclerosis progression. Scientifica (Cairo) 2016: 8514056.
- Horn P, Baars T, Kahlert P, Heiss C, Westenfeld R, et al. (2015) Release of intracoronary microparticles during stent implantation into stable atherosclerotic lesions under protection with an aspiration device. PLoS One 10: e0124904.
- Morel O, Luca F, Grunebaum L, Jesel L, Meyer N, et al. (2011) Short-term very low-calorie diet in obese females improves the haemostatic balance through the reduction of leptin levels, PAI-1 concentrations and a diminished release of platelet and leukocyte-derived microparticles. Int J Obes 35: 1479-1486.
- Liu Y, He Z, Zhang Y, Dong Z, Bi Y, et al. (2016) Dissimilarity of increased phosphatidylserine-positive microparticles and associated coagulation activation in acute coronary syndromes. Coron Artery Dis.
- Suades R, Padró T, Vilahur G, Martin-Yuste V, Sabaté M, et al. (2015) Growing thrombi release increased levels of CD235a(+) microparticles and decreased levels of activated platelet-derived microparticles. Validation in ST-elevation myocardial infarction patients. J Thromb Haemost 13: 1776-1786.
- Suades R, Padró T, Crespo J, Ramaiola I, Martin-Yuste V, et al. (2016) Circulating microparticle signature in coronary and peripheral blood of ST elevation myocardial infarction patients in relation to pain-to-PCI elapsed time. Int J Cardiol 202: 378-387.
- Sansone R, Stanske B, Keymel S, Schuler D, Horn P, et al. (2015) Macrovascular and microvascular function after implantation of left ventricular assist devices in end-stage heart failure: Role of microparticles. J Heart Lung Transplant 34: 921-932.
- Empana JP, Boulanger CM, Tafflet M, Renard JM, Leroyer AS, et al. (2015) Microparticles and sudden cardiac death due to coronary occlusion. The TIDE (Thrombus and Inflammation in sudden Death) study. Eur Heart J Acute Cardiovasc Care 4: 28-36.
- Wang ZT, Wang Z, Hu YW (2016) Possible roles of platelet-derived microparticles in atherosclerosis. Atherosclerosis 248: 10-16.
- Montoro-García S, Shantsila E, Tapp LD, López-Cuenca A, Romero AI, et al. (2013) Small-size circulating microparticles in acute coronary syndromes: relevance to fibrinolytic status, reparative markers and outcomes. Atherosclerosis 227: 313-322.
- Morel O, Pereira B, Averous G, Faure A, Jesel L, et al. (2009) Increased levels of procoagulant tissue factor-bearing microparticles within the occluded coronary artery of patients with ST-segment elevation myocardial infarction: role of endothelial damage and leukocyte activation. Atherosclerosis 204: 636-641.
- Petrini S, Neri T, Lombardi S, Cordazzo C, Balìa C, et al. (2016) Leptin induces the generation of procoagulant, tissue factor bearing microparticles by human peripheral blood mononuclear cells. Biochim Biophys Acta 1860: 1354-1361.
- Burnouf T, Goubran HA, Chou ML, Devos D, Radosevic M (2014) Platelet microparticles: detection and assessment of their paradoxical functional roles in disease and regenerative medicine. Blood Rev 28: 155-166.
- Osumi K, Ozeki Y, Saito S, Nagamura Y, Ito H, et al. (2001) Development and assessment of enzyme immunoassay for platelet-derived microparticles. Thromb Haemost 85: 326-330.
- Tan KT, Lip GY (2005) The potential role of platelet microparticles in atherosclerosis. Thromb Haemost 94: 488-492.
- Suades R, Padró T, Alonso R, Mata P, Badimon L (2015) High levels of TSP1+/CD142+ platelet-derived microparticles characterise young patients with high cardiovascular risk and subclinical atherosclerosis. Thromb Haemost 114: 1310-1321.
- Van der Zee PM, Biró E, Ko Y, de Winter RJ, Hack CE, et al. (2006) P-selectin- and CD63-exposing platelet microparticles reflect platelet activation in peripheral arterial disease and myocardial infarction. Clin Chem 52: 657-664.
- Min PK, Kim JY, Chung KH, Lee BK, Cho M, et al. (2013) Local increase in microparticles from the aspirate of culprit coronary arteries in patients with ST-segment elevation myocardial infarction. Atherosclerosis 227:323-328.
- Suades R, Padró T, Vilahur G, Badimon L (2012) Circulating and platelet-derived microparticles in human blood enhance thrombosis on atherosclerotic plaques. Thromb Haemost 108: 1208-1219.
- Michelsen AE, Brodin E, Brosstad F, Hansen JB (2008) Increased level of platelet microparticles in survivors of myocardial infarction. Scand J Clin Lab Invest 68: 386-389.
- Alexandru N, Andrei E, Dragan E, Georgescu A (2015) Interaction of platelets with endothelial progenitor cells in the experimental atherosclerosis: Role of transplanted endothelial progenitor cells and platelet microparticles. Biol Cell 107: 189-204.
- Berezin AE (2015) Are endothelial cell-derived microparticles predictive biomarkers in cardiovascular diseases? Atherosclerosis 1: e101-e103.
- Mause SF, Ritzel E, Liehn EA, Hristov M, Bidzhekov K, et al. (2010) Platelet microparticles enhance the vasoregenerative potential of angiogenic early outgrowth cells after vascular injury. Circulation 122: 495-506.
- Baj-Krzyworzeka M, Majka M, Pratico D, Ratajczak J, Vilaire G, et al. (2002) Platelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cells. Exp Hematol 30: 450-459.
- Ohtsuka M, Sasaki K, Ueno T, Seki R, Nakayoshi T, et al. (2013) Platelet-derived microparticles augment the adhesion and neovascularization capacities of circulating angiogenic cells obtained from atherosclerotic patients. Atherosclerosis 227: 275-282.
- Chen BA, Zhong YJ, Huang CY, Li CP, Shi GY, et al. (2007) Effects of platelet-derived membrane microparticles on angiogenesis in chick chorioallantoic membranes. Zhongguo Shi Yan Xue Ye Xue Za Zhi 15: 1070-1073.
- Sinning JM, Losch J, Walenta K, Böhm M, Nickenig G, et al. (2011) Circulating CD31+/Annexin V+ microparticles correlate with cardiovascular outcomes. Eur Heart J 32: 2034-2041.
- Werner N, Wassmann S, Ahlers P, Kosiol S, Nickenig G (2006) Circulating CD31+/annexin V+ apoptotic microparticles correlate with coronary endothelial function in patients with coronary artery disease. Arterioscler Thromb Vasc Biol 26: 112-116.
- Huang PH, Huang SS, Chen YH, Lin CP, Chiang KH, et al. (2010) Increased circulating CD31+/annexin V+ apoptotic microparticles and decreased circulating endothelial progenitor cell levels in hypertensive patients with microalbuminuria. J Hypertens 28: 1655-1665.
- Porto I, Biasucci LM, De Maria GL, Leone AM, Niccoli G, et al. (2012) Intracoronary microparticles and microvascular obstruction in patients with ST elevation myocardial infarction undergoing primary percutaneous intervention. Eur Heart J 33: 2928-2938.
- Berezin AE, Kremzer AA, Samura TA, Berezina TA, Kruzliak P (2015) Immune phenotypes of endothelial-derived microparticles in dysmetabolic patients. J Proteomics Bioinformatics 8: 060-066.
- Chironi GN, Boulanger CM, Simon A, Dignat-George F, Freyssinet JM, et al. (2009) Endothelial microparticles in diseases. Cell Tissue Res 335: 143-151.
- Shantsila E (2009) Endothelial microparticles: a universal marker of vascular health? J Hum Hypertens 23: 359-361.
- Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V, et al. (2011) Exosome/microvesicle mediated epigenetic reprogramming of cells. Am J Cancer Research 1: 98-110.
- Théry C, Ostrowski M, Segura E (2009) Membrane vesicles as conveyors of immune responses. Nat Rev Immunol 9: 581-593.
- Wang Y, Chen LM, Liu ML (2014) Microvesicles and diabetic complications - novel mediators, potential biomarkers and therapeutic targets. Acta Pharmacol Sin 35: 433-443.
- Mause SF, Weber C (2010) Microparticles: protagonists of a novel communication network for intercellular information exchange. Circ Res 107: 1047-1057.
- Montoro-García S, Shantsila E, Marín F, Blann A, Lip GY (2011) Circulating microparticles: new insights into the biochemical basis of microparticle release and activity. Basic Res Cardiol 106: 911-923.
- Berezin AE (2015) Circulating endothelial-derived apoptotic microparticles as novel perspective biomarker for diabetes. Diabetes Res Treat Open Access 1: 117-120.
- Berezin AE, Kremzer AA, Martovitskaya YV, Berezina TA, Gromenko EA (2016) Pattern of endothelial progenitor cells and apoptotic endothelial cell-derived microparticles in chronic heart failure patients with preserved and reduced left ventricular ejection fraction. E Bio Med 4: 86-94.
- Berezin AE, Kremzer AA, Samura TA, Berezina TA, Kruzliak P (2015) Impaired immune phenotype of circulating endothelial-derived microparticles in patients with metabolic syndrome and diabetes mellitus. J Endocrin Inves 38: 865-874.
- Berezin A, Kremzer A, Cammarota G, Zulli A, Petrovic D, et al. (2015) Circulating endothelial-derived apoptotic microparticles and insulin resistance in non-diabetic patients with chronic heart failure. Clin Chem Lab Med.
- Berezin AE, Kremzer AA, Martovitskaya YUV, Berezina TA, Samura TA (2015) Pattern of circulating endothelial-derived microparticles among chronic heart failure patients with dysmetabolic comorbidities: The impact of subclinical hypothyroidism. Diab Metab Syn Clin Res Rev 10: 29-36.
- Berezin AE, Kremzer AA, Samura TA, Berezina TA (2015) Immune phenotype of circulating endothelial-derived microparticlesin elderly patients with metabolic syndrome and diabetes mellitus. J Gerontol Geriatr Res 4: 199-209.
- Berezin A (2015) Small-size endothelial-derived microparticles as novel predictive biomarkers in cardiovascular diseases. Angiol 3: 145-148.
- Bank IE, Timmers L, Gijsberts CM, Zhang YN, Mosterd A, et al. (2015) The diagnostic and prognostic potential of plasma extracellular vesicles for cardiovascular disease. Expert Rev Mol Diag 15: 1577-1588.
- Berezin AE, Kremzer AA, Berezina TA, Martovitskaya YUV (2015) Pattern of circulating microparticles in chronic heart failure patients with metabolic syndrome: Relevance to neurohumoral and inflammatory activation. BBA Clinical 4: 69-75.
- Berezin AE, Kremzer AA, Martovitskaya YV, Samura TA, Berezina TA (2014) The predictive role of circulating microparticles in patients with chronic heart failure. BBA Clin 3: 18-24.
- Sen S, McDonald SP, Coates PT, Bonder CS (2011) Endothelial progenitor cells: novel biomarker and promising cell therapy for cardiovascular disease. Clin Sci 120: 263-283
- Amabile N, Heiss C, Chang V, Angeli FS, Damon L, et al. (2009) Increased CD62e(+) endothelial microparticle levels predict poor outcome in pulmonary hypertension patients. J Heart Lung Transplant 28: 1081-1086.
- Shantsila E, Lip GY (2016) Endothelial progenitors and blood microparticles: are they relevant to heart failure with preserved ejection fraction? EBioMedicine 4: 5-6.
- Berezin AE, Kremzer AA, Samura TA, Martovitskaya YV, Malinovskiy YV, et al. (2015) Predictive value of apoptotic microparticles to mononuclear progenitor cells ratio in advanced chronic heart failure patients. J Cardiol 65: 403-411.
- Berezin AE (2015) Predicting heart failure phenotypes using cardiac biomarkers: hype and hope. J Dis Markers 2: 1035-1041.
- Berezin A, Kremzer A, Martovitskaya Y, Samura T, Berezina T (2016) The novel biomarker risk prediction score in patients with chronic heart failure. Clin Hypertension 22: 1.
- Berezin A (2015) Impaired phenotype of circulating endothelial-derived microparticles: novel marker of cardiovascular risk. J Cardiol Ther 2: 273-278.
- Berezin AE (2016) Prognostication in different heart failure phenotypes: the role of circulating biomarkers. J Circulating Biomarkers 5: 01.