Keywords
Alcohol abuse; HIV; AIDS; ER; Golgi apparatus; Liver disorders
Introduction
Globally, there are 36.7 million people living with HIV and approximately 1.8 million acquire HIV infection every year [1]. In the United States, 1.2 million people are living with HIV infection and half million with AIDS have died since the HIV epidemic began in the 1980’s [1-3]. The situation demands development of antiretroviral drugs and access to antiretroviral therapies. While anti-HIV vaccines or genome editing are yet to be developed and put into practice, antiretroviral agents are effective [4-6]. For instance, HIV protease inhibitors (HIV PIs) that inhibit HIV proteinase or protease are used in the highly active effective antiretroviral therapies (HAART) and nucleic acid-based anti-HIV compounds are developed for further inhibition of intracellular viral targets. The quality of life of HIV/AIDS patients under anti- HIV therapies is improved significantly. However, there are numerous reports indicating that some antivirals singly or in combination increase the risk of comorbidities [7-9]. While some side effects of anti-HIV drugs are manageable, some can be very serious and fatal. For instance, hepatic injuries have emerged as the major non-AIDS–related cause of death among HIV/ AIDS patients [8,10]. The risk of drug side effects gets worse as nearly half of the HIV-infected patients abuse/consume alcohol, which not only impairs patients’ adherence to HAART but also worsens anti-HIV drug-induced hepatotoxicity leading to greater morbidity and mortality [11,12]. Nonhazardous alcohol use of less than five standard drinks (equivalent to 5 × 14 g of pure alcohol) once a week can reduce survival of HIV/AIDS patients by one year and daily hazardous use of five or more standard drinks per day reduces survival by more than six years [11-13]. Increase in liver injuries such as liver fibrosis or cirrhosis are often seen at all levels of alcohol exposure [8,10,11]. To be even complicated, there are 10 million HIV/AIDS patients worldwide co-infected with hepatitis viruses and/or tuberculosis (TB). Additional drugs against the co-infections and/or drug abuse could further increase the severity of the hepatoxicities [8,14,15]. Therefore, it is of importance to dissect pathogenic mechanisms underlying the hepatoxicity caused by the drugs combined with alcohol consumption/abuse, which would provide basis for a better management of AIDS patients suffering from liver disease.
Organelle Stress in Antivirals and Alcohol-Induced Liver Disorders
There are a few potential mechanisms underlying the alcohol and drug-induced hepatotoxicity/liver injury: (1) direct intrinsic toxicities from individual drugs [16]; (2) idiosyncratic hypersensitivity reactions [16]; (3) aberrant immune activities [16,17]; (4) metabolic abnormality and cellular stress response [18]. The intrinsic toxicity occurs dose-dependently at sub-lethal doses and can be influenced by environmental and genetic sensitivity factors. The idiosyncratic reactions occur in a minority of patients without obvious relationship to drug dose or time of onset. The innate and adaptive immune responses are generally known to be involved in the liver toxicity. Detailed characteristics of these potential mechanisms are not discussed in this focused review. The stress and metabolic abnormality mechanism is the most significant and relevant as cellular stress responses in the liver may generate danger signals which co-stimulate the immune system, and alcohol and majority of the anti-HIV drugs are metabolized in the liver by the cytochrome P450 enzyme system, which is bound to interfere with the metabolism and the antiviral therapies in AIDS patients. In fact, alcohol as “the first hit” is reported to affect hepatic CYP activities that metabolize protease inhibitors [18-20]. Both alcohol and certain antiviral drugs induce organelle stress, such as the induction of unfolded protein accumulation in the endoplasmic reticulum (ER) resulting in ER stress and cell death [20,21]. Anti-HIV drugs such as ritonavir (RTV), indinavir (IDV), lopinavir (LPV) or atazanavir (ATZ) have been reported to induce ER stress in hepatic cells [22]. We initially discovered that alcohol induced ER stress in the liver of animal models [23]. The ER stress normally triggers protective unfolded protein response (UPR), which involves three ER stress sensors: IRE1 (inositol requiring enzyme 1), PERK (PKR-like ER kinase) and ATF6 (activating transcription factor), to restore ER homeostasis and minimize injuries. However, prolonged UPR such as under conditions of chronic alcohol consumption and/or long-term anti-HIV therapies induces fat accumulation, inflammation and cell death, which are well documented to lead to development of hepatic steatosis, fibrosis and cirrhosis [20-25]. Because the ER stress response is a major pathogenesis mechanism, xenobiotics to ensure proper ER homeostasis have been developed and tested in a variety of in vitro and in vivo model systems. They include: molecular chaperones such as sodium 4-phenylbutyrate (PBA) and taurine conjugated ursodeoxycholic acid (TUDCA) that increase the ER protein-folding capacity; and compounds such as antioxidants, autophagy inducers, and UPR enhancers (e.g. salubrinal/guana benz, trans-4,5-dihydroxy-1,2-dithiane (DTTox), and valproate) that either increase expression of protein chaperones or enhance the protective UPR pathways [21,25,26]. However, the xenobiotics for ER homeostasis restoration resulted in partial protective effects, suggesting that although the ER stress is involved in the alcohol and drug-induced liver injuries, there are other cellular targets that either contribute or are upstream of the ER stress response.
The organelle that is closely associated with the ER is the Golgi apparatus. The Golgi is part of the cellular endomembrane system, in which secretory and membrane proteins from the ER receive various modifications such as glycosylation, phosphorylation and sulfation and are then packaged into membrane-bound vesicles before trafficking to their destinations. Similar to the ER, the structure and capacity of the Golgi can fluctuate according to physiological demands or pathological stress conditions. When the protein load and modifications exceed the Golgi capacity there is a Golgi stress response (referred to as GSR) to increase its capacity [20,27]. Impaired GSR can cause cellular injuries directly or indirectly through the ER stress. There are a few factors that may regulate GSR. The first is TFE3 (a basic-helix-loop-helix type transcription factor) [28], which regulates transcriptional activation of genes encoding vesicular trafficking components or Golgi residents such as RAB20, syntaxin 3A (STX3A), protein 60 (GCP60), GM130, giantin, sialyltransferase, fucosyltransferase and glycosylation enzymes. Translocation of TFE3 into the nucleus depends on the status of its phosphorylation. Upon Golgi stress, TFE3 is dephosphorylated and translocated into the nucleus where it activates the GSR genes. The second is the CREB3-ARF4 pathway involving both ER and Golgi [29]. CREB3 is a basic leucine zipper-containing transcription factor that resides in the ER membrane. ARF4 (ADP-ribosylation factor 4) is a member of the small GTPase family that regulates Golgi-to-ER vesicular trafficking. CREB3 is activated via proteolysis and upregulates the transcription of ARF4 in Brefeldin A (BFA) treated cells that are under both ER and Golgi stresses. BFA is known to inhibit the function of several guanine nucleotide exchange factors (GEFs) and blocks GEF-mediated Golgi-to-ER trafficking [30]. The third pathway involves an ER chaperone, HSP47. HSP47 may protect cells from the Golgi stress as expression of HSP47 is increased under the Golgi stress and suppression of HSP47 by siRNA resulted in fragmentation of the Golgi apparatus and cell death [31].
Alcohol consumption/abuse has long been known to induce ultrastructural changes in the Golgi. In man and animal, chronic alcohol feeding with nutritionally adequate diets induced ultrastructural abnormalities of the intestinal epithelial cells, mammary cells, hepatocytes, neurons, and glandular epithelium cells [32-38]. In addition to the morphological changes, there is evidence for alcohol-induced functional and metabolic changes. Chronic alcohol exposure affects the ER-Golgi trafficking in neuronal dendrites [39]. Alcohol alters glycosyl transferase activity in the Golgi of liver cells [40]. Acute ethanol intoxication interferes with various steps of protein glycosylation at the Golgi of rat liver [41]. Further, accumulations of lipid and carbohydrates and decreased terminal glycosylation were observed in the Golgi of isolated hepatocytes treated with alcohol, which was associated with production of anticytoplasmic autoantibodies. Significantly, high titers of anti-Golgi antibodies were detected in human alcoholics with hepatocellular carcinoma (HCC) [42].
There is also evidence that either viral infection or antiviral drugs induce the Golgi stress. For instance, hepatitis C (HCV) replication requires the guanine nucleotide exchange factor 1 (GBF1) and its effector ADP ribosylation factor 1 (Arf1) that are known to regulate Golgi membrane trafficking and organelle structure in the secretory pathway. HCV infection is reported to result in Golgi fragmentations [43]. On the other hand, pegylated interferon (Peg-IFN) plus ribavirin, the standard therapy for HCV, induces anti-Golgi antibodies, which is associated with liver injury in patients [44].
Emerging Evidence and Hypothesis of Disrupted ER-Golgi Trafficking in Drugs and Alcohol-Induced Liver Pathogenesis
Synergistic or additive effects of antivirals combined with alcohol on liver injury have been observed recently [8,18,45,46]. Chronic alcohol feeding of animal models resulted in fat accumulation, which is associated with distorted morphologies of the ER and Golgi organelles visible under electron microscopy (Figure 1). Treating the animals with a standard regimen for HIV-infected patients (i.e., ritonavir boosted lopinavir) induced moderate dilatation of the ER and dispersed Golgi apparatus. Combination treatments with alcohol and the drugs induced severe damages to the hepatocytes. Neither the ER nor the Golgi was identifiable and rare myelin-like structures surrounding giant lipid droplets were observed for the first time in the liver cells from treated animals. In contrast, the hepatocytes from control animals had lamella ER and normal Golgi apparatus.
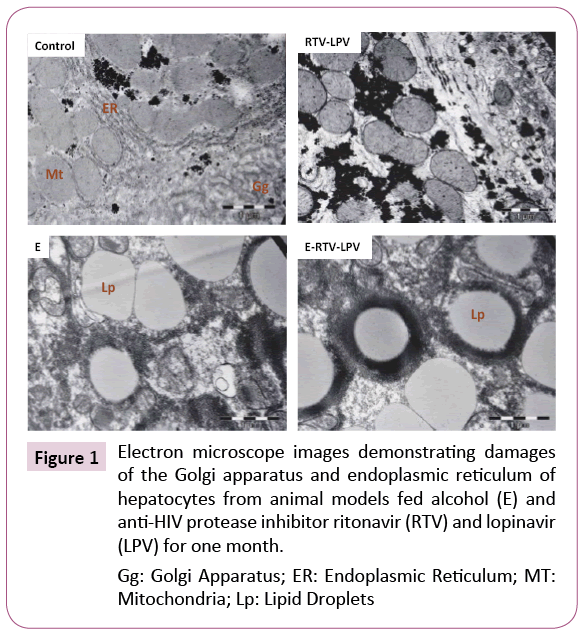
Figure 1: Electron microscope images demonstrating damages of the Golgi apparatus and endoplasmic reticulum of hepatocytes from animal models fed alcohol (E) and anti-HIV protease inhibitor ritonavir (RTV) and lopinavir (LPV) for one month.
Gg: Golgi Apparatus; ER: Endoplasmic Reticulum; MT: Mitochondria; Lp: Lipid Droplets
In pursuing molecular details of the alcohol and drug-induced organelle stress response, we found that the three canonical UPR branches, IRE, PERK and ATF6 were differentially expressed in HepG2 or primary hepatocytes in response to the RTV boosted LPV treatment [18,46]. The ATF6 branch as well as genes/factors downstream of the ATF6 were inactivated or not altered while the other two branches of UPR were upregulated. These observations are of great interest as the activation of ATF6 is known to require ER-to-Golgi trafficking and proteolytic processing that involves both the ER and the Golgi apparatus [47]. In fact, co-localization of ATF6 and the Golgi was lower in the liver cells treated with the drugs and alcohol than in the cells treated with the specific ER stress inducing agent tunicamycin or thapsigargin [46]. In parallel to the reduction of ATF6 in the Golgi, marked Golgi fragmentation was observed in the RTV and LPV-treated liver cells (Figure 2), which was concentration and time dependent. Of note, the Golgi fragmentation was not due to mitosis as the drug-induced fragmentation was detected in both HepG2 cells and rarely dividing primary hepatocytes and liver tissues [46]. Apoptosis did not cause the fragmentation either as increased caspase activities were not detected until hours after the drug treatments and pancaspase inhibitors did not show any rescue effects on the druginduced Golgi fragmentation. Moreover, variations in severity of the fragmentation were observed in response to other anti-HIV drugs including amprenavir, darunavir and nelfinavir, which were correlated with downstream ER stress and cell death and fatty liver injury [46]. All these pieces of evidence suggest that Golgi dysfunction and disturbed ER-Golgi trafficking contributes to the anti-HIV drugs and/or alcohol-induced liver disorders.
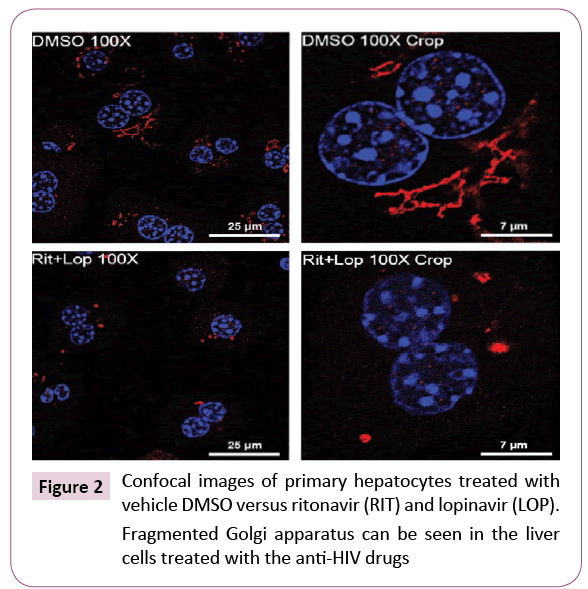
Figure 2: Confocal images of primary hepatocytes treated with vehicle DMSO versus ritonavir (RIT) and lopinavir (LOP).
Fragmented Golgi apparatus can be seen in the liver cells treated with the anti-HIV drugs
The ER-Golgi trafficking is bi-directional and responsible for biogenesis and intracellular distribution of biomolecules [48]. ER-to-Golgi trafficking or anterograde transportation mediated by the COPII complexes moves newly synthesized proteins and lipids to Golgi for processing, sorting and redistribution. Meanwhile, Golgi-to-ER trafficking or retrograde transportation mediated by the COPI complexes ensures recycling of lipids, fluids and ER escaped proteins [48,49]. Impairments of either anterograde or retrograde could eventually collapse the whole ER-Golgi trafficking and trigger cellular stress responses leading to injury [20,21,46,48]. Thus, integrity of the ER-Golgi trafficking is essential for maintaining Golgi morphology [48-50]. The Golgi fragmentation observed by us indicates that the drugs stress the Golgi and disrupt the ER-Golgi trafficking. Indeed, Golgi stress response markers, GCP60 and HSP47 were increased in the drugtreated liver cells and knockdown of TFE3 worsened the druginduced cell death [46]. Further, the anterograde ER-to-Golgi trafficking is likely affected more than the retrograde trafficking by the anti-HIV drugs and alcohol. This is because the effects of the drugs on Golgi fragmentation are similar to the effects of H89. H89 is a protein kinase A (PKA) inhibitor. PKA is required for the initiation and assembly of the COPII complexes [51]. In addition, BFA is known to inhibit specifically the assembly of COPI complexes [30] and its effects on the Golgi fragmentation are different from the effects of the anti-HIV drugs. Thus, it is conceivable that anti-HIV drugs interfere with the ER-Golgi trafficking and Golgi integrity causes Golgi stress and/or ER stress (Figure 3). The Golgi stress triggers Golgi stress response, which either restores Golgi homeostasis or induces injury depending on stress conditions. Alcohol consumption may deteriorate the drug-induced Golgi dysfunction and hepatic injury through interfering with the metabolism of the anti-HIV drugs as five to ten-fold increases of the drug concentrations were observed in the blood of animals fed alcohol [18,46].
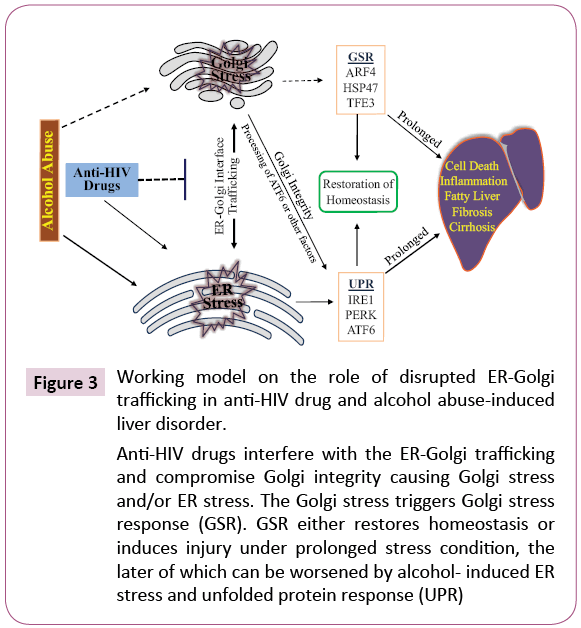
Figure 3: Working model on the role of disrupted ER-Golgi trafficking in anti-HIV drug and alcohol abuse-induced liver disorder.
Anti-HIV drugs interfere with the ER-Golgi trafficking and compromise Golgi integrity causing Golgi stress and/or ER stress. The Golgi stress triggers Golgi stress response (GSR). GSR either restores homeostasis or induces injury under prolonged stress condition, the later of which can be worsened by alcohol- induced ER stress and unfolded protein response (UPR)
Perspectives
The primary target of antiviral drug and alcohol abuse appears to be the ER-Golgi trafficking based on our recent observations, which could explain a few aspects of pathological consequences in the hepatocytes that synthesize and secrete large amounts of substances. Disruptions in the ER-Golgi trafficking will have impact on the integrity and function of the Golgi apparatus. The stressed Golgi either respond to restore its homeostasis that potentially involves ARF4, HSP47 and TFE3 or dissemble to generate danger signals to stimulate the general immune response that eliminates the cells containing the damaged Golgi. The impaired ER-Golgi trafficking could also suspend processing and activation of transcription factors such as ATF6 that is involved in maintaining ER homeostasis. Loss of the ER homeostasis will stress the ER which is well established to induce cell death and disease development. Thus, vicious cycles resulted from the disrupted ER-Golgi trafficking most likely occurs in the liver cells that metabolize both the drugs and alcohol.
The question is what specific molecular components of the ERGolgi traffic machineries are affected by the anti-HIV drugs and/ or alcohol. The trafficking between ER and Golgi mediated by the endomembrane system involves vesicle budding, uncoating, docking and fusion that are often through recruitments of effectors such as vesicle tethers, SNAREs, membrane and motor proteins. These effectors are regulated by the Rab GTPases that belong to a large family of small GTP-binding proteins and accomplish their functions by switching between an inactive GDP bound and an active GTP-bound form. About one third of the 60 members of the Rab GTPase family found in human cells have been associated with either the ER or Golgi complex or the membrane intermediates at their interface. Since majority of the Rab proteins and effectors are not well characterized, it is hard to predict specific targets of the antiviral drugs. However, considering that the drugs were initially developed to prevent viral replication by selectively binding to viral proteases (e.g. HIV-1 protease) and blocking proteolytic cleavage of protein precursors required for the virus production, one would speculate that the drugs could also have some unintended effects on certain Rab proteins or effectors of the ER-Golgi trafficking machinery that require proteolytic cleavage for their maturation or functional activation. Future research should be directed to seek these unintended targets, which would provide molecular basis for modifications of the current anti-HIV drugs so that minimize the side effects of anti-HIV or other virus medicine.
Acknowledgement
Supported by grants from the U.S. National Institute of Drug Abuse (R01 DA042632) and National Institute on Alcohol Abuse and Alcoholism (R21 AA023952).
References
- UNAIDS Fact Sheet Latest statistics on the status of the AIDS epidemic (2016).
- Piot P, Quinn TC (2013) Response to the AIDS pandemic--a global health model. N Engl J Med 368: 2210-2218.
- Centers for Disease Control and Prevention (2016) Twenty-five years of HIV/AIDS–United States, 1981-2006. MMWR Morb Mortal Wkly Rep 55: 585-589.
- Baeten J, Celum C (2013) Systemic and topical drugs for the prevention of HIV infection: Antiretroviral pre-exposure prophylaxis. Annu Rev Med 64: 219-232.
- Palesch D, Kirchhoff F (2013) First steps toward a globally effective HIV/AIDS vaccine. Cell 155: 495-497.
- Kwarteng A, Ahuno ST, Kwakye-Nuako G (2017) The therapeutic landscape of HIV-1 via genome editing. AIDS Res Ther 14: 32.
- Kumar S, Rao PS, Earla R, Kumar A (2015) Drug-drug interactions between anti-retroviral therapies and drugs of abuse in HIV systems. Expert Opin Drug Metab Toxicol 11: 343-355.
- Sherman KE, Rockstroh J, Thomas D (2015) Human immunodeficiency virus and liver disease: An update. Hepatology 62: 1871-1882.
- Menshawy A, Ismail A, Abushouk AI, Ahmed H, Menshawy E, et al. (2017) Efficacy and safety of atazanavir/ritonavir-based antiretroviral therapy for HIV-1 infected subjects: A systematic review and meta-analysis. Arch Virol 162: 2181-2190.
- Acharya C, Dharel N, Sterling RK (2015) Chronic liver disease in the human immunodeficiency virus patient. Clin Liver Dis 19: 1-22.
- US National Institute on Alcohol Abuse and Alcoholism (2015) Alcohol facts and statistics.
- Williams EC, Hahn JA, Saitz R, Bryant K, Lira MC, et al. (2016) Alcohol use and human immunodeficiency virus (HIV) infection: Current knowledge, implications and future directions. Alcohol Clin Exp Res 40: 2056-2072.
- Braithwaite RS, Conigliaro J, Roberts MS, Shechter S, Schaefer A, et al. (2007) Estimating the impact of alcohol consumption on survival for HIV+ individuals. AIDS Care 19: 459-466.
- Compton WM, Volkow ND (2006) Major increases in opioid analgesic abuse in the United States: Concerns and strategies. Drug Alcohol Depend 81: 103-107.
- Petoumenos K, Law MG (2016) Smoking, alcohol and illicit drug use effects on survival in HIV-positive persons. Curr Opin HIV AIDS 11: 514-520.
- Kaplowitz N (2005) Idiosyncratic drug hepatotoxicity. Nat Rev Drug Discov 4: 489-499.
- Sulkowski MS (2003) Hepatotoxicity associated with antiretroviral therapy containing HIV-1 protease inhibitors. Semin Liver Dis 23: 183-194.
- Kao E, Shinohara M, Feng M, Lau MY, Ji C (2012) Human immunodeficiency virus protease inhibitors modulate Ca2+ homeostasis and potentiate alcoholic stress and injury in mice and primary mouse and human hepatocytes. Hepatology 56: 594-604.
- Kumar S, Jin M, Ande A, Sinha N, Silverstein PS, et al. (2012) Alcohol consumption effect on antiretroviral therapy and HIV-1 pathogenesis: Role of cytochrome P450 isozymes. Expert Opin Drug Metab Toxicol 8: 1363-1375.
- Ji C (2015) Advances and new concepts in alcohol-induced organelle stress, unfolded protein responses and organ damage. Biomolecules 5: 1099-1121.
- Ji C, Kaplowitz N, Han H (2017) Role of endoplasmic reticulum stress in hepatic injury. In: Ding WX, Yin XM (eds) Molecules, systems and signaling in liver injury. Cell Death in Biology and Diseases, Springer Cham 221-250.
- Zhou H, Gurley EC, Jarujaron S, Ding H, Fang Y, et al. (2006) HIV protease inhibitors activate the unfolded protein response and disrupt lipid metabolism in primary hepatocytes. Am J Physiol Gastrointest Liver Physiol 291: G1071-G1080.
- Ji C, Kaplowitz N (2003) Betaine decreases hyperhomocysteinemia, endoplasmic reticulum (ER) stress and liver injury in alcohol fed mice. Gastroenterology 124: 1488-1499.
- Ji C, Kaplowitz N, Lau MY, Kao E, Petrovic LM, et al. (2011) Liver-specific loss of glucose-regulated protein 78 perturbs the unfolded protein response and exacerbates a spectrum of liver diseases in mice. Hepatology 54: 229-239.
- Cao SS, Kaufman RJ (2013) Targeting endoplasmic reticulum stress in metabolic disease. Expert Opin Ther Targets 17: 437-448
- Lafleur MA, Stevens JL, Lawrence JW (2013) Xenobiotic perturbation of ER stress and the unfolded protein response. Toxicol Pathol 41: 235-262.
- Taniguchi M, Yoshida H (2017) TFE3, HSP47 and CREB3 pathways of the mammalian Golgi stress response. Cell Struct Funct 42: 27-36.
- Taniguchi M, Nadanaka S, Tanakura S, Sawaguchi S, Midori S, et al. (2015) TFE3 is a bHLH-ZIP-type transcription factor that regulates the mammalian Golgi stress response. Cell Struct Funct 40: 13-30.
- Reiling JH, Olive AJ, Sanyal S, Carette JE, Brummelkamp TR, et al. (2013) A CREB3-ARF4 signaling pathway mediates the response to Golgi stress and susceptibility to pathogens. Nat Cell Biol 15: 1473-1485.
- Helms JB, Rothman JE (1992) Inhibition by brefeldin A of a Golgi membrane enzyme that catalyses exchange of guanine nucleotide bound to ARF. Nature 360: 352-354.
- Miyata S, Mizuno T, Koyama Y, Katayama T, Tohyama M (2013) The endoplasmic reticulum-resident chaperone heat shock protein 47 protects the Golgi apparatus from the effects of O-glycosylation inhibition. PLoS ONE 8: e69732.
- Rubin E, Rybak BJ, Lindenbaum J, Gerson CD, Walker G, et al. (1972) Ultra structural changes in the small intestine induced by ethanol. Gastroenterology 63: 801-814.
- Vilaró S, Viñas O, Remesar X (1989) Altered ultrastructure of lactating rat mammary epithelial cells induced by chronic ethanol ingestion. Alcohol Clin Exp Res 13: 128-136.
- Matsuda Y, Takada A, Takase S, Yasuhara M (1991) Effects of ethanol on the secretion of hepatic secretory protein in rat alcoholic liver injury. Alcohol 8: 433-437.
- Petrosyan A, Cheng PW, Clemens DL, Casey CA (2015) Down regulation of the small GTPase SAR1A: A key event underlying alcohol-induced Golgi fragmentation in hepatocytes. Sci Rep 5: 17127.
- Madeira MD, Sousa N, Lieberman AR, Paula-Barbosa MM (1993) Effects of chronic alcohol consumption and of dehydration on the supraoptic nucleus of adult male and female rats. Neuroscience 56: 657-672.
- Sousa N, Madeira MD, Ruela C, Paula-Barbosa MM (1995) Structural reorganization in the supraoptic nucleus of withdrawn rats following long-term alcohol consumption. Alcohol Clin Exp Res 19: 879-885.
- Cagnon VH, Tomazini FM, Garcia PJ, Martinez M, Padovani CR, et al. (2001) Structure and ultrastructure of the ventral prostate of isogenic mice (C57B1/6J) submitted to chronic alcohol ingestion. Tissue Cell 33: 354-360.
- Romero AM, Renau-Piqueras J, Marín MP, Esteban-Pretel G (2015) Chronic alcohol exposure affects the cell components involved in membrane traffic in neuronal dendrites. Neurotox Res 27: 43-54.
- Gang H, Lieber CS, Rubin E (1973) Ethanol increases glycosyl transferase activity in the hepatic Golgi apparatus. Nat New Biol 243: 123-125.
- Cottalasso D, Gazzo P, Dapino D, Domenicotti C, Pronzato MA, et al. (1996) Effect of chronic ethanol consumption on glycosylation processes in rat liver microsomes and Golgi apparatus. Alcohol Alcohol 31: 51-59.
- Mozo L, Simó A, Suárez A, Rodrigo L, Gutiérrez C (2002) Autoantibodies to Golgi proteins in hepatocellular carcinoma: Case report and literature review. Eur J Gastroenterol Hepatol 14: 771-714.
- Hansen MD, Johnsen IB, Stiberg KA, Sherstova T, Wakita T, et al. (2017) Hepatitis C virus triggers Golgi fragmentation and autophagy through the immunity-related GTPase M. Proc Natl Acad Sci USA 114: E3462-E3471.
- Paraná R, Schinoni MI, de Freitas LA, Codes L, Cruz M, et al. (2006) Anti-Golgi complex antibodies during pegylated-interferon therapy for hepatitis C. Liver Int 26: 1148-1154.
- Hu J, Han H, Lau MY, Lee H, MacVeigh-Aloni M, et al. (2015) Effects of combined alcohol and anti-HIV drugs on cellular stress responses in primary hepatocytes and hepatic stellate and kupffer cells. Alcohol Clin Exp Res 39: 11-20.
- Han H, He Y, Hu J, Lau R, Lee H, et al. (2017) Disrupted ER-to-Golgi trafficking underlies anti-HIV drugs and alcohol-induced cellular stress and hepatic injury. Hepatol Commun 1: 122-139.
- Ye J, Rawson RB, Komuro R, Chen X, Davé UP, et al. (2000) ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol Cell 6: 1355-1364.
- Brandizzi F, Barlowe C (2013) Organization of the ER-Golgi interfaces for membrane traffic control. Nat Rev Mol Cell Biol 14: 382-392.
- Borgese N (2016) Getting membrane proteins on and off the shuttle bus between the endoplasmic reticulum and the Golgi complex. J Cell Sci 129: 1537-1545.
- Sengupta P, Satpute-Krishnan P, Seo AY, Burnette DT, Patterson GH, et al. (2015) ER trapping reveals Golgi enzymes continually revisit the ER through a recycling pathway that controls Golgi organization. PNAS 112: E6752-E6761.
- Lee TH, Linstedt AD (2000) Potential role for protein kinases in regulation of bidirectional endoplasmic reticulum-to-Golgi transport revealed by protein kinase inhibitor H89. Mol Biol Cell 11: 2577-2590.